
Динозавры — Википедия
Диноза́вры[1](лат. Dinosauria) — надотряд пресмыкающихся из клады Dracohors, возникший в верхнетриасовую эпоху, между 243 и 233,23 млн лет назад. Динозавры стали доминирующими наземными позвоночными после триасово-юрского вымирания 201 млн лет назад; их доминирование продолжалось в течение юрского и мелового периодов. Палеонтологическая летопись показывает, что птицы — это современные пернатые динозавры, эволюционировавшие от более ранних теропод в конце юрского периода. Условно всех динозавров, за исключением птиц, называют нептичьими динозаврами.
Динозавры — разнообразная группа животных с таксономической, морфологической и экологической точек зрения. Птицы, насчитывающие &&&&&&&&&&010770.&&&&&010 770 современных видов и более 20 000 подвидов[2], являются самой разнообразной группой позвоночных после окунеобразных. Используя ископаемые свидетельства, палеонтологи выявили более 500 различных родов
В то время как предки динозавров были двуногими (бипедальными), многие вымершие группы динозавров включали в себя четвероногие виды, а некоторые из них были способны менять эти позиции. Сложные демонстрационные структуры, такие как рога и гребни, являются общими для всех групп динозавров, а некоторые вымершие представители развили такие модификации, как костяная броня и шипы. В то время как современные выжившие птичьи динозавры (птицы), как правило, невелики из-за ограничений размера, необходимого для полёта, многие доисторические динозавры (как нептичьи, так и птичьи) были крупнотелыми: самые большие динозавры-завроподы по оценкам достигали длины в 39,7 м при высоте в 18 м и были самыми крупными наземными животными всех времён. Тем не менее идея, что все нептичьи динозавры были гигантскими, является ошибочным представлением, основанным частично на том, что вероятность разрушения больших и прочных костей до того, как они окаменеют, гораздо меньше, чем вероятность того, что то же самое произойдёт с костями маленького животного. Многие нептичьи динозавры были довольно маленькими: например,
С тех пор, как первые окаменелые кости динозавров были опознаны и признаны в начале 19 века, их ископаемые скелеты стали главными достопримечательностями музеев по всему миру, а динозавры превратились в неотъемлемую часть популярной культуры. Большие размеры некоторых групп динозавров, а также распространившийся чудовищный и фантастический образ, обеспечили регулярное появление динозавров в бестселлерах и фильмах, таких как «Парк юрского периода». Постоянный общественный энтузиазм к этим животным привел к значительному финансированию палеонтологии, и новые открытия регулярно освещаются в средствах массовой информации.
Термин «динозавр» ввёл в 1842 году английский биолог Ричард Оуэн для описания первых ископаемых остатков древних ящеров, поражавших воображение учёных своими размерами[6]. Слово происходит от др.-греч. δεινός [1](

Согласно филогенетической номенклатуре, динозавры обычно определяются как группа, состоящая из самого последнего общего предка (MRCA) трицератопса и птиц, а также всех его потомков. Также предлагалось, чтобы динозавры определялись в отношении MRCA мегалозавра и игуанодона, поскольку это два из трёх родов, отнесённых Ричардом Оуэном к динозаврам в статье, в которой он выделил данную группу.
Птицы в настоящее время признаны единственной сохранившейся линией динозавров-теропод. В традиционной систематике птицы считались отдельным классом, произошедшим от динозавров, отдельного надотряда. Однако большинство современных палеонтологов, изучающих динозавров, отвергают традиционную систематику в пользу филогенетической таксономии. Этот подход предполагает, что для того, чтобы группа была естественной, все потомки членов группы также должны быть включены в неё. Таким образом, птицы считаются динозаврами и, следовательно, динозавры не вымерли
Исследования Мэттью Барона, Дэвида Нормана и Пола Баретта 2017-го года предположили радикальный пересмотр систематики динозавров, который пока что не получил общепризнанности. Филогенетический анализ Барона и коллег восстановил птицетазовых как более близких к тероподам, чем к завроподоморфам, как это считается традиционно. Они восстановили кладу орнитосцелиды, чтобы дать название группе, объединяющей птицетазовых и теропод. Сами динозавры были определены как общий предок грубого трицератопса, домового воробья и диплодока Карнеги и все его потомки, чтобы гарантировать, что завроподы и их родственники будут однозначно подходить под определение динозавров

Общее описание[править | править код]
Используя одно из приведённых выше определений, динозавров можно в целом описать как архозавров с задними конечностями, удерживаемыми прямо под телом. Зачастую динозаврами ошибочно называют животных, на самом деле к роду динозавров нисколько не относящихся. Так, например, летающими динозаврами называют птерозавров, хотя данный термин корректнее было бы использовать по отношению к большинству представителей клады авиал, то есть к птицам и их близким родственникам, морскими динозаврами по ошибке называют ихтиозавров, плезиозавров и мозазавров. Зачастую непосвящённые люди относят к динозаврам ранних синапсидов. Также в культуре распространено ложное мнение, что каждый «завр» — это динозавр, хотя, на самом деле, данное окончание довольно часто используется и в названиях других животных. Так, например, выделяется род доисторических китообразных базилозавр.
Динозавры были доминирующими наземными позвоночными мезозоя, особенно в юрский и меловой периоды. Остальные группы животных, за исключением некоторых других архозавров (например, наземных крокодиломорфов), были сильно ограничены в размерах и биологических нишах. Млекопитающие, например, редко превышали размера современной кошки, и, как правило, становились добычей плотоядных динозавров
Динозавры были чрезвычайно разнообразной группой животных, по данным исследования 2006 года, на тот момент были с уверенностью описаны более 500 родов нептичьих динозавров, а общее число родов, указанных в палеонтологической летописи, по оценкам, могло достигать около 1850, около 75% из которых ещё не обнаружено
В 2016 году количество предполагаемое количество видов динозавров, существовавших в мезозойскую эру, было оценено в 1543—2468[17][18]. Некоторые из них были травоядными, в том числе употреблявшими в пищу семена, другие плотоядными и всеядными. В то время как некоторые динозавры (в том числе птицы) передвигались на двух конечностях, некоторые группы нептичьих динозавров ходили на четырёх, а некоторые их представители, такие как
Отличительные анатомические особенности[править | править код]
Хотя недавние открытия затруднили представление общепризнанного списка отличительных черт динозавров, почти все динозавры, описанные до сих пор, разделяют определённые модификации скелета, присущие ещё нединозавровым архозаврам или более древним архозавроморфам, от которых те произошли. Хотя некоторые более поздние группы динозавров отличались несколько другими, дальнейшими модификациями этих признаков, они всё-таки считаются типичными для динозавров; самые ранние динозавры имели их и передали их своим потомкам. Такие модификации, происходящие от самого последнего общего предка некоторой таксономической группы, называются синапоморфиями такой группы[21].
Детальная оценка взаимоотношений архозавров Стерлинга Несбитта подтвердила или обнаружила следующие двенадцать однозначных синапоморфий, некоторые из которых были известны ранее[22]:
- В черепе надвисочная ямка присутствует перед надвисочным окном, главным отверстием в заднем дерматокрауме
- Эпипофизы, косо направленные назад отростки на задних верхних углах, присутствуют в передних шейных позвонках позади атланта и эпистрофея, второго шейного позвонка
- Лучевая кость, нижняя кость передней конечности, короче плечевой кости на 80%
- Четвертичный трохантер (область, в которой каудофеморальная мышца прикрепляется к внутреннему заднему валу) на бедренной кости представляет собой острый выступ
- Четвертичный трохантер ассиметричен, обладает дистальным нижним краевым образованием более крутого угла к валу
- На таранной и пяточной костях, верхних костях голеностопного сустава, проксимальная суставная фасетка, верхняя соединительная поверхность, на малоберцовой кости занимает менее 30% поперечной ширины элемента
- Внезатылочные кости (кости в задней части черепа) отсутствуют вдоль средней линии на нижней части эндокринной полости, внутреннего пространства мозговой оболочки
- В области таза проксимальные суставные поверхности седалищной кости с подвздошной и лобковой костями отделены большой вогнутой поверхностью (на верхней стороне седалищной кости часть открытого тазобедренного сустава расположена между контактами с лобковой и подвздошной костями)
- Поясничный гребень большеберцовой кости, выступающая часть верхней поверхности большеберцовой кости, дугообразно изгибается спереди и снаружи
- Отчетливый проксимодистально ориентированный (вертикальный) гребень присутствует на задней грани дистального конца большеберцовой кости (задняя поверхность нижнего конца большеберцовой кости)
- Вогнутая суставная поверхность малоберцовой кости пяточной кости (верхняя поверхность пяточной кости, где она соприкасается с малоберцовой костью, имеет полый контур)
Несбитт представил ряд дальнейших потенциальных синапоморфий и отбросил ряд синапоморфий, предложенных ранее. Некоторые из них также присутствуют у силезаврид, которых Несбитт восстановил в качестве сестринской группы динозавров, в том числе большой передний трохантер, плюсневые кости II и IV субэквивалентной длины, уменьшенный контакт между седалищной и лобковой костями, наличие кнемиального креста на голени и восходящего процесса на астрагале и многие другие[23].
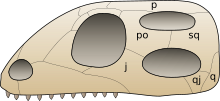
Множество других скелетных особенностей характерны для всех динозавров. Однако, поскольку они присущи и другим группам архозавров, либо отсутствовали у некоторых ранних динозавров, эти особенности не считаются синапоморфиями. Например, как диапсиды, имеют две пары височных окон (отверстий в черепе за глазами) и как члены диапсидной группы архозавров, имеют дополнительные отверстия в передней части черепа и нижней челюсти[24]. Кроме того, известно, что некоторые характеристики, которые раньше считались синапоморфиями, появились до появления динозавров или отсутствовали у самых ранних динозавров и независимо развились у различных групп динозавров. К ним относятся удлиненная лопатка; крестец, состоящий из трёх или более слитых позвонков (три встречаются у некоторых других архозавров, но у герреразавра их только два)[23] и перфорация вертлужной впадины или тазобедренного сустава с отверстием в её центре внутренней поверхности (к примеру, закрыт у сатурналии)[25][26]. Другая трудность определения чётко выраженных особенностей динозавров состоит в том, что ранние динозавры и другие архозавры позднего триаса часто плохо известны и во многих отношениях были похожи; эти животные иногда неправильно идентифицировались в литературе[27].
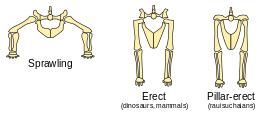 Тазобедренные суставы и позы задних конечностей (слева направо): типичных пресмыкающихся (раскидистые), динозавров и млекопитающих (прямые) и равизухий (прямые)
Тазобедренные суставы и позы задних конечностей (слева направо): типичных пресмыкающихся (раскидистые), динозавров и млекопитающих (прямые) и равизухий (прямые)Задние конечности динозавров, как у большинства современных млекопитающих, выпрямлены, но отличаются от таких у большинства других пресмыкающихся, чьи конечности растягиваются по обе стороны[28]. Это положение обусловлено развитием углубления в области таза, обращённого вбок (обычно открытого углубления и соответствующей обращённой внутрь головки на бедре[29]). Их вертикальное положение позволяло ранним динозаврам легко дышать во время движения, что, вероятно, способствовало выносливости и уровням активности, которые превышали таковые у «ползучих» пресмыкающихся[30]. Вертикальные конечности, вероятно, также помогали поддержать развитие большого размера, уменьшая изгибающие напряжения на конечностях. У некоторых нединозавровых архозавров, включая равизухий, также были прямые конечности, но это было достигнуто с помощью конфигурации тазобедренного сустава, в которой вместо выступа бедренной вставки на углублении на бедре, верхняя тазовая кость поворачивалась, формируя нависающий выступ[31].
Предки[править | править код]
Рептилии обитали на Земле ещё до появления динозавров. Лапы у них были расположены по бокам туловища, как у большинства современных ящериц. Около 300 млн лет назад во время каменноугольного периода произошло глобальное потепление, что, по мнению учёных, уничтожило тропические леса и вызвало эволюционный взрыв среди рептилий. Каждая популяция оказалась запертой в небольшой области обитания, и каждая начала развиваться по-своему, что и привело к увеличению разнообразия[32].
Затем появились предки динозавров — архозавры («господствующие ящеры»). Появление архозавров шло параллельно с массовым (но не полным) вымиранием терапсид на границе перми и триаса (см. Массовое пермское вымирание). В начале триасового периода возникло множество новых видов.
Важнейшие продвинутые признаки ранних динозавров, которые отсутствовали у большинства текодонтов, были связаны с положением тела и характером локомоции. Задние конечности становятся практически вертикальными, а их длина по сравнению с более короткими передними конечностями позволяет делать выводы о хождении на двух ногах (бипедальности)[33].
Высшие формы терапсид по своей организации были очень близки к однопроходным млекопитающим и, по некоторым предположениям[34], имели молочные железы и шерсть. Внезапный упадок тероморфов и расцвет «настоящих ящеров» — одна из самых интригующих загадок палеонтологии[35].
Динозавры отклонились от своих предков архозавров спустя примерно 20 миллионов лет после массового пермского вымирания, которое уничтожило приблизительно 95 % всей жизни на Земле[36]. На сегодняшний день остатки самых древних динозавров имеют возраст около 230 миллионов лет. Однако в 2010 году группа американских, немецких и южноафриканских палеонтологов представила в научном мире совместную работу, согласно которой первые динозавры населяли Землю на несколько миллионов лет раньше, чем это было принято считать. Свои выводы они сделали, проанализировав остатки новой динозаврообразной рептилии Asilisaurus kongwу, которая является родственным динозаврам видом, жившим 245—240 млн лет назад. Учёные говорят, что уже в это время различия между Dinosauriformes и самыми ранними динозаврами были существенными, следовательно, общий предок динозавров отделился от группы орнитодир значительно раньше, чем считалось прежде[37].
По одной из гипотез, оба отряда динозавров впервые появились на территории современной Южной Америки и затем распространились по всей Пангее в течение триасового периода с 230 до 199 миллионов лет назад[38][39].
Первые динозавры[править | править код]
 Скелеты эрреразавра и эораптора
Скелеты эрреразавра и эораптораОдним из самых ранних известных на сегодняшний день динозавров считается ставрикозавр, живший 228 млн лет назад, частичный скелет которого был обнаружен в отложениях среднего триасового периода на территории современной Бразилии. Большая берцовая кость длиннее бедренной, что характерно для бегающих животных, кости конечностей полые[40]. В 1963 году был описан эрреразавр, обнаруженный в отложениях позднего триаса на северо-западе Аргентины. Из-за фрагментарности остатков точная его классификация была не определена. Лишь после того как в 1988 году был обнаружен более полный скелет с черепом, он в основном стал классифицироваться как примитивный теропод[41]. Обнаруженный на территории современной Мексики вид Tawa hallae подтвердил связь эрреразавра с более поздними тероподами (хищными динозаврами)[42][43]. Эрреразавр достигал длины до 4 метров[40] и веса 200—250 килограммов[41]. Череп длинный и узкий, не имеет признаков, характерных для более поздних теропод[41], строение задних конечностей также характерно для ранних динозавров[40]. Через четыре года после открытия эрреразавра, в Аргентине был найден скелет ещё одного древнего динозавра — эораптора[44], жившего 228—235 млн лет назад. Внешне он был похож на других рептилий, но особенности костей и зубов позволяют отнести его к примитивным тероподам[45]. Палеонтологи полагают, что эораптор напоминает общего предка всех динозавров[46]. Если эти предположения верны, то основываясь на его чертах, можно говорить, что первые динозавры были маленькими, двуногими хищниками[44]. Открытия примитивных, подобных динозаврам орнитодиров, таких как маразух и Lagerpeton, поддерживают эти предположения; анализ окаменелостей данных рептилий действительно говорит об их малых размерах и хождении на двух ногах.
Самой первой известной группой птицетазовых динозавров являются Fabrosauridae (фаброзавриды). Самый ранний из них — пизанозавр (Pisanosaurus), обнаруженный в породах верхнего триаса на территории современной Аргентины. Лучше всего из фаброзавридов изучен род лесотозавр из юрских отложений Южной Африки. Как и все другие фаброзавриды, он достигал в длину около 1 метра. Строение задних конечностей характерно для бипедальных животных, большая берцовая кость значительно длиннее бедренной. Некоторые части скелета сходны с таковыми у ранних теропод, но тазовые кости и строение черепа и зубов уже имеют признаки птицетазовых динозавров[40].
На самых ранних этапах эволюции динозавров земная среда была заселена различными видами архозавров и терапсидов, такими как этозавры, цинодонты, дицинодонты, орнитозухиды, ринхозавры и другие. По всей видимости, в конце триаса динозавры выдержали долгую конкуренцию с данными животными, прежде чем победили и достигли расцвета. Некоторые учёные, однако, полагают, что господствующее положение динозавров было получено не столько превосходством над другими животными, сколько случайностью. В карнийском веке динозавры не показывают образец устойчивого увеличения в разнообразии и числе, как то должно было иметь место, если бы они, конкурируя, занимали ниши других групп животных. Вместо этого они были очень редки, составляя только 1—2 % от общей животной фауны. Лишь в норийском веке, после исчезновения нескольких других групп, они стали существенными компонентами фауны, представляя уже 50—90% живых существ[47]. Около 200 миллионов лет назад произошло так называемое триасовое вымирание. Это событие окончательно освободило экологические ниши, открыв возможность развиться, начиная с юрского периода, таким группам, как крокодиломорфы, птерозавры, млекопитающие, черепахи и динозавры[48].
Диверсификация в меловом периоде[править | править код]
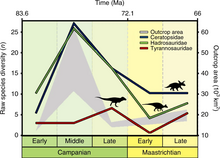
Статистические исследования, основанные на имеющихся палеонтологических материалах, предполагают, что разнообразие динозавров, то есть число разновидностей, увеличилось в позднем меловом периоде.
Однако в июле 2008 года Грэм Т. Ллойд и его коллеги выдвинули утверждение, что это очевидное разнообразие является иллюзией, вызванной тем, что породы позднего мелового периода изучены в большей степени, чем породы юрского и триасового периодов. Вместо этого они написали статью о том, что в позднем меловом периоде существенной диверсификации подверглись только две группы динозавров — гадрозавры и цератопсы. В среднем меловом периоде цветущие покрытосеменные растения стали большей частью наземных экосистем, в то время как ранее доминировали голосеменные, такие как хвойные породы. Копролиты (окаменелый навоз) динозавров указывают на то, что в то время как некоторые ели покрытосеменные растения, большинство растительноядных динозавров питалось, главным образом, голосеменными растениями. В это же время растительноядные насекомые и млекопитающие развивались очень быстро и разносторонне, чтобы использовать в своих интересах новый тип пищи растительного происхождения. Ящерицы, змеи, крокодилы и птицы также разносторонне развивались в то же самое время. Ллойд и его коллеги предположили, что отказ нептичьих динозавров от разностороннего развития в свете изменения экосистемы обрекал их на исчезновение[49].
Динозавры относятся к группе архозавров, или «господствующих рептилий». К этой группе также относятся крокодилы, птерозавры и их родственники.
В филогенетической систематике динозавры обычно определяются как группа, состоящая из «трицератопсов, неорнитов (современных птиц), их общих предков и всех потомков»[48]. Также некоторыми учёными выдвигается предложение, чтобы динозавры были определены относительно общего предка мегалозавра и игуанодона, так как они были двумя из трёх первых родов, процитированных Ричардом Оуэном, когда он впервые выделил группу динозавров[50].
Динозавры делятся на отряды ящеротазовых и птицетазовых по форме своего таза. Такое деление было предложено в 1887 году британским палеонтологом Гарри Сили (Harry Seeley). Названия носят условный характер, так как у ящеротазовых динозавров строение таза вовсе не похоже на строение его у ящериц, а птицы происходят не от птицетазовых, а от ящеротазовых динозавров. Ящеротазовые — отряд динозавров с лобковыми костями, первично направленными вперед (как у крокодилов). У некоторых относительно поздних ящеротазовых лобковые кости становятся направленными назад (как у птиц). У птицетазовых лобковые кости направлены назад изначально.
Значение этих различий не ясно. Среди обеих групп имеются как четвероногие формы, так и двуногие, однако именно по этим особенностям определяется принадлежность динозавров к той или другой группе, независимо от их размеров, способа передвижения и питания.
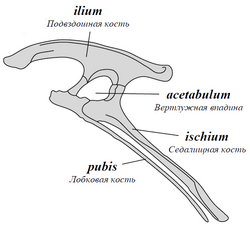 | |
| Тазовые кости птицетазовых динозавров. Лобковые кости направлены назад | Тазовые кости ящеротазовых динозавров. Лобковые кости выступают вперёд |
Более объяснимы различия между группами в строении челюстей и зубов. У ящеротазовых динозавров зубы располагаются по краю челюсти в один ряд, который доходит до конца морды. Каждый зуб конической или долотообразной формы сидит в отдельной ячейке. У птицетазовых динозавров в передней части нижней челюсти располагалась лишённая зубов предзубная кость, часто отсутствовали передние зубы и в верхней челюсти. У многих птицетазовых был роговой клюв, как у черепах.
К ящеротазовым относят тероподов («звероногие») и завроподоморф («ящероногих»). Подотряд тероподы включает в себя всех хищных ящеров, из которых наиболее крупным и известным является тираннозавр. Считается, что от мелких тероподов произошли птицы. Завроподоморфы — в основном крупные динозавры с длинными шеями и хвостами и колоннообразными ногами. Они делятся на зауропод (брахиозавры, апатозавры, диплодоки и т. п.) и прозауропод, представлявших побочную ветвь зауропод).
Птицетазовые включает следующие группы: стегозавры («пластинчатые ящеры»), у которых пластины расположены вдоль позвоночника; анкилозавры («окостенелые ящеры»), покрытые костяной броней по всему телу; цератопсы («рогатые ящеры»), наиболее известен трицератопс; пахицефалозавры («толстоголовые ящеры») с головами, покрытыми костяным панцирями, и орнитоподы («птиценогие»), для которых характерно передвижение преимущественно на двух ногах (бипедальность), их наиболее известный представитель — игуанодон.
Согласно традиционной классификации, стегозавров и анкилозавров объединяют в подотряд тиреофоров («щитоносных»), цератопсов и пахицелозавров — в подотряд маргиноцефалов («бахромчатоголовых»), и третьим подотрядом считают орнитоподов. По новейшей классификации выделяют два подотряда: тиреофоров и цераподов, а в последний относят орнитоподов, цератопсов и пахицефалозавров в качестве инфраотрядов.
Альтернативная классификация[править | править код]
Есть и другие гипотезы разделения надотряда[51]:
Мел-палеогеновое вымирание — Википедия
Миллионов лет назад
Мел-палеогеновое вымирание (мел-третичное, мел-кайнозойское, K-T вымирание) — одно из пяти т. н. «великих массовых вымираний», на границе мелового и палеогенового периода, около 66 миллионов лет назад. Частью этого массового вымирания явилось вымирание нептичьих динозавров[1].
Вместе с нептичьими динозаврами вымерли прогрессивные морские завропсиды, в том числе мозазавры и плезиозавры, летающие ящеры (птерозавры), многие моллюски, в том числе аммониты и белемниты, и множество мелких водорослей. Всего погибло 16 % семейств морских животных (47 % родов морских животных) и 18 % семейств сухопутных позвоночных, включая практически всех крупных и средних по размеру. Все существовавшие в мезозое экосистемы были полностью разрушены, что впоследствии резко подстегнуло эволюцию таких групп животных, как птицы и млекопитающие, давших в начале палеогена огромное многообразие форм благодаря освобождению большинства экологических ниш[2][3].
Тем не менее, большинство таксономических групп растений и животных на уровнях от отряда и выше пережило этот период. Так, не вымерли мелкие сухопутные завропсиды, такие как змеи, черепахи, ящерицы и птицы, а также крокодиломорфы, включая доживших до наших дней крокодилов. Выжили ближайшие родственники аммонитов — наутилусы, млекопитающие, кораллы и наземные растения.
Существует предположение, что некоторые нептичьи динозавры (гадрозавры, тероподы и др.) существовали на западе Северной Америки и в Индии в течение ещё нескольких миллионов лет в начале палеогена после их вымирания в других местах (палеоценовые динозавры[en])[4]. Причём это предположение плохо согласуется с любым из сценариев импактного вымирания[2].
| Докембрий | Фанерозой | Эон | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Палеозой | Мезозой | Кайнозой | Эра | ||||||||||
| Кембрий | Ордо вик | Сил ур | Девон | Карбон | Пермь | Триас | Юра | Мел | Палео ген | Нео ген | П-д | ||
| 4570 | 541 | 485,4 | 443,4 | 419,2 | 358,9 | 298,9 | 252,2 | 201,3 | 145,0 | 66,0 | 23,03 | млн лет ← | |
| 2,588 | |||||||||||||
На конец 1990-х годов ещё не существовало единой точки зрения на причину и характер данного вымирания[5][6].
К середине 2010-х годов дальнейшие исследования данного вопроса привели к тому, что в научном сообществе возобладала точка зрения, гласящая, что важной причиной мел-палеогенового вымирания было падение небесного тела, вызвавшее появление кратера Чиксулуб на полуострове Юкатан, иные точки зрения рассматривались в качестве маргинальных[7][2][8]. В настоящее время эта точка зрения не была опровергнута, но было предложено множество иных, альтернативных или дополняющих факторов, которые также могли сыграть свою роль в массовом вымирании.
 | Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
Внеземные гипотезы[править | править код]
- Гипотеза импакта. Падение астероида — одна из самых распространённых версий[9] (так называемая «гипотеза Альвареса», обнаружившего мел-палеогеновую границу). Она основана главным образом на приблизительном соответствии времени образования кратера Чиксулуб (который является следом от падения метеорита размером порядка 10 км около 65 млн лет назад[10]) на полуострове Юкатан в Мексике и временем вымирания большинства из исчезнувших видов динозавров[11]. Кроме того, небесно-механические расчёты (основанные на наблюдениях ныне существующих астероидов) показывают, что метеориты размером более 10 км сталкиваются с Землёй в среднем около одного раза в 100 млн лет, что по порядку величины соответствует, с одной стороны, датировкам известных кратеров, оставленных такими метеоритами[12], а с другой — промежуткам времени между пиками вымираний биологических видов в фанерозое[13]. Теорию подтверждает повышенное содержание иридия и других платиноидов в тонком слое на границе известняковых отложений мела и палеогена, отмеченное во многих районах мира. Эти элементы имеют тенденцию концентрироваться в мантии и ядре Земли и очень редко встречаются в поверхностном слое. С другой стороны, химический состав астероидов и комет точнее отражает первоначальное состояние Солнечной системы, в котором иридий занимает более существенное положение[14][15]. С помощью компьютерного моделирования учёные показали, что в воздух было выброшено около 15 трлн тонн пепла и сажи, и днём на Земле было темно, как лунной ночью. В результате нехватки света у растений замедлился[16] или на 1—2 года был ингибирован[17]фотосинтез, что могло привести к уменьшению концентрации кислорода в атмосфере (на время, пока Земля была закрыта от поступления солнечного света). Температура на континентах упала на 28 °C, в океанах — на 11 °C. Исчезновение фитопланктона, важнейшего элемента пищевой цепи в океане, привело к вымиранию зоопланктона и других морских животных[17]. В зависимости от времени пребывания в стратосфере сульфатных аэрозолей глобальная годовая средняя температура приземного воздуха уменьшалась на 26 °C, до 16 лет температура была ниже +3 °C[18]. Залегающий между толщей суевита[en] или импактной брекчии и вышележащим палеоценовым пелагическим известняком 76-сантиметровый переходный слой в кратере Чиксулуб, включая верхнюю часть со следами ползания и рытья (en:Trace fossil), сформировался менее, чем за 6 лет после падения астероида[19]. В пользу гипотезы, объясняющей вымирание падением небесного тела, свидетельствует геологически мгновенное повышение уровня кислотности поверхностного слоя океана на мел-палеогеновой границе (снижение pH составило 0,2—0,3), выявленное при изучении изотопов бора в известковых оболочках ископаемых фораминифер. До этого момента уровень кислотности был стабильным в течение последних 100 тысяч лет мелового периода. За резким повышением кислотности последовал период постепенного нарастания щёлочности (повышение pH на 0,5), продолжавшийся до 40 тысяч лет от мел-палеогеновой границы. Возвращение кислотности к первоначальному уровню заняло ещё 80 тысяч лет. Такие явления могут быть объяснены снижением потребления щелочей в связи с вымиранием кальцинирующего планктона из-за быстрого закисления поверхностных вод ливневым выпадением SO2 и NOx, попавших в атмосферу в результате удара крупного болида[20].
- Версия «многократного падения» (англ. multiple impact event), предполагающая несколько последовательных ударов[10]. Она привлекается, в частности, для объяснения того, что вымирание произошло не одномоментно (смотри раздел Недостатки гипотез). Косвенно в её пользу свидетельствует тот факт, что метеорит, создавший кратер Чиксулуб, был одним из осколков более крупного небесного тела[21]. Некоторые геологи считают, что кратер Шива на дне Индийского океана, датируемый примерно тем же временем, является следом падения второго гигантского метеорита, ещё большего[22], но эта точка зрения является дискуссионной. Существует компромисс между гипотезами удара одного или нескольких метеоритов — столкновение с двойной системой метеоритов. Параметры кратера Чиксулуб подходят для такого удара, если оба метеорита были меньше, но вместе имели примерно те же размер и массу, что и метеорит гипотезы одного столкновения[23].
- Взрыв сверхновой звезды либо близкий гамма-всплеск.
- Столкновение Земли с кометой. Именно этот вариант рассматривается в сериале «Прогулки с динозаврами». Известный американский физик Лиза Рэндалл связывает гипотезу падения кометы на Землю с влиянием темной материи[24].
 Столкновения Земли с астероидом: художественное изображение
Столкновения Земли с астероидом: художественное изображениеЗемные абиотические[править | править код]
- Усиление вулканической активности[25], с которой связывают ряд эффектов, которые могли бы повлиять на биосферу: изменение газового состава атмосферы; парниковый эффект, вызванный выбросом углекислого газа при извержениях; изменение освещённости Земли из-за выбросов вулканического пепла (вулканическая зима). В пользу этой гипотезы говорят геологические свидетельства о гигантском излиянии магмы в промежутке между 68 и 60 млн лет назад на территории Индостана, в результате которого образовались деканские траппы[26].
- Резкое понижение уровня моря, произошедшее в последней (маастрихтской) фазе мелового периода («маастрихтская регрессия»).
- Изменение среднегодовых и сезонных температур. Это было бы особенно актуально в случае валидности предположения об инерциальной гомойотермии крупных динозавров, которая требовала бы ровного тёплого климата[27]. Вымирание, однако, не совпадает по времени со значительным изменением климата[28], и, согласно современным исследованиям, динозавры были скорее полностью теплокровными животными (см. физиология динозавров).
- Резкий скачок магнитного поля Земли.
- Переизбыток кислорода в атмосфере Земли.
- Резкое охлаждение океана.
- Изменение состава морской воды.
Земные биотические[править | править код]
- Эпизоотия — массовая эпидемия.
- Динозавры не смогли приспособиться к изменению типа растительности и отравились алкалоидами, содержащимися в появившихся цветковых растениях (с которыми, однако, сосуществовали в течение десятков миллионов лет, причём именно с появлением цветковых растений был связан эволюционный успех отдельных групп травоядных динозавров, освоивших новый биом травянистых степей).
- На численность динозавров сильно повлияли первые хищные млекопитающие, уничтожая кладки яиц[en] и детёнышей.
- Вариация предыдущей версии о вытеснении нептичьих динозавров млекопитающими. Между тем, все меловые млекопитающие — очень мелкие, в основном насекомоядные животные. В отличие от завропсидов, которые благодаря целому ряду прогрессивных специализаций, включая появление чешуи и перьев, яиц в плотной оболочке и живорождения, сумели в своё время освоить принципиально новую среду — удалённые от водоёмов сухие ландшафты, млекопитающие не имели никаких принципиальных эволюционных преимуществ по сравнению с современными им рептилиями. Метаболизм как минимум некоторых динозавров был столь же интенсивен, как у млекопитающих, на что указывают изотопные, сравнительно-морфологические, гистологические и географические данные. Следует отметить, что очень сложно отличить наиболее обособившихся манирапторов от примитивных птиц, эти группы имели отличия на уровне семейств и отрядов, а не классов; в кладистике они рассматриваются как разные отряды одного класса завропсидов.
- Иногда всплывает гипотеза, что часть крупных морских рептилий могла не выдержать конкуренции с появившимися именно в это время акулами современного типа. Однако ещё в девоне акулы зарекомендовали себя как неконкурентоспособная в отношении более высокоразвитых позвоночных группа, будучи отодвинутыми на задний план костными рыбами. Очень крупные и довольно прогрессивные на фоне своих сородичей акулы возникали в позднем меловом периоде после упадка плезиозавров, но они быстро были вытеснены начавшими занимать освободившиеся ниши мозазаврами.
«Биосферная» версия[править | править код]
В русской палеонтологии популярна биосферная версия «великого вымирания», в том числе вымирания нептичьих динозавров[27]. Необходимо отметить, что большинство из выдвинувших её палеонтологов специализировались на изучении не динозавров, а других животных: млекопитающих, насекомых, и так далее. Согласно ей, основными исходными факторами, предопределившими исчезновение нептичьих динозавров и других крупных пресмыкающихся, стали:
- Появление цветковых растений.
- Постепенное изменение климата, вызванное дрейфом материков.
Последовательность событий, приведшая к вымиранию, представляется следующим образом:
- Цветковые растения, имеющие более развитую корневую систему и лучше использующие плодородие почвы, достаточно быстро повсеместно вытеснили прочие виды растительности. При этом появились насекомые, специализированные на питании цветковыми, а насекомые, «привязанные» к ранее существовавшим видам растительности, начали вымирать.
- Цветковые растения образуют дернину, являющуюся лучшим из природных подавителей эрозии. В результате их распространения снизилось размывание поверхности суши и, соответственно, поступление в океаны питательных веществ. «Обеднение» океана пищей привело к гибели значительной части водорослей, являвшихся основным первичным производителем биомассы в океане. По цепочке это привело к полному нарушению всей морской экосистемы и стало причиной массовых вымираний в море. Это же вымирание затронуло и крупных летающих ящеров, которые, по имеющимся представлениям, были трофически связаны с морем.
- На суше животные активно приспосабливались к питанию зелёной массой (кстати, и травоядные динозавры тоже). В малом размерном классе появились мелкие фитофаги-млекопитающие (типа современных крыс). Их появление привело к появлению и соответствующих хищников, которыми тоже стали млекопитающие. Малоразмерные хищники-млекопитающие были неопасны для взрослых динозавров, но питались их яйцами и детёнышами, создавая динозаврам дополнительные трудности в воспроизводстве. При этом охрана потомства для крупных динозавра практически неосуществима из-за слишком большой разницы в размерах взрослых особей и детёнышей.
 | Легко наладить охрану кладки (некоторые динозавры в позднем мелу действительно отрабатывают такие типы поведения), однако когда детёныш имеет размер кролика, а родители — ростом со слона, то его быстрее раздавишь, чем защитишь от нападения.[27] |  |
- Из-за жёсткого ограничения на максимальный размер яйца (обусловленного допустимой толщиной скорлупы) у крупных видов динозавров детёныши рождались намного более лёгкими, чем взрослые особи (у самых крупных видов разница в массе между взрослыми и детёнышами составляла тысячи раз). А это означает, что все крупные динозавры в процессе роста должны были неоднократно менять свою пищевую нишу, причём на ранних этапах развития им приходилось конкурировать с более специализированными в определённых размерных классах видами[29]. Отсутствие передачи опыта между поколениями только усугубляло данную проблему.
- В результате дрейфа материков в конце мелового периода изменилась система воздушных и морских течений, что привело к некоторому похолоданию на значительной части суши и усилению сезонного температурного градиента, что заметно отразилось на биосфере. Динозавры, будучи специализированной группой, были наиболее уязвимы для таких изменений. Динозавры не были теплокровными животными, и само изменение температуры могло послужить значимым фактором их вымирания.
В результате всех перечисленных причин для нептичьих динозавров создались неблагоприятные условия, которые и привели к прекращению появления новых видов. «Старые» виды динозавров ещё некоторое время существовали, но постепенно вымерли полностью. Судя по всему, жёсткой прямой конкуренции динозавров и млекопитающих не было, они занимали разные размерные классы, существуя параллельно. Лишь после исчезновения динозавров млекопитающие захватили освободившуюся экологическую нишу, да и то не сразу.
Что любопытно, развитие первых архозавров в триасовом периоде сопровождалось постепенным вымиранием многих терапсид, высшие формы которых являлись по сути примитивными яйцекладущими млекопитающими[27].
Комбинированные[править | править код]
Вышеперечисленные гипотезы могут дополнять друг друга, что некоторыми исследователями используется для выдвижения разного рода комбинированных гипотез. Например, удар гигантского метеорита мог спровоцировать усиление вулканической активности и выброс большой массы пыли и пепла, что в совокупности могло повлечь за собой изменение климата, а это, в свою очередь — изменение типа растительности и пищевых цепочек, и т. д.; изменение климата также могло быть вызвано понижением уровня Мирового океана. Деканские вулканы начали извергаться ещё до падения метеорита, однако в определённый момент частые и мелкие извержения (71 тысяча кубометров в год) сменились редкими и масштабными (900 миллионов кубометров в год). Учёные допускают, что смена типа извержений могла произойти под влиянием упавшего в то же самое время метеорита (с погрешностью в 50 тысяч лет)[30][31].
Известно, что у некоторых рептилий наблюдается явление зависимости пола потомства от температуры кладки яиц. В 2004 году группа исследователей из британского Университета Лидса, которую возглавляет Дэвид Миллеангл. David Miller), предположила, что если подобное явление было характерно и для динозавров, то изменение климата всего на несколько градусов могло спровоцировать появление на свет особей только определённого пола (мужского, например), а это, в свою очередь, делает невозможным дальнейшее размножение[32].
Недостатки гипотез[править | править код]
Ни одна из перечисленных гипотез не может в полной мере объяснить весь комплекс явлений, связанных с вымиранием нептичьих динозавров и других видов в конце мелового периода.
Главные проблемы перечисленных версий следующие:
- Гипотезы фокусируются именно на вымирании, которое, как считает часть исследователей, шло теми же темпами, что и в предшествующее время, но при этом перестали образовываться новые виды в составе вымерших групп.
- Все импактные гипотезы (гипотезы ударного воздействия), в том числе астрономические, не соответствуют предполагаемой продолжительности его периода (многие группы животных начали вымирать задолго до конца мела, и есть доказательства существования палеогеновых динозавров, мозазавров и других животных). Переход тех же аммонитов к гетероморфным формам тоже свидетельствует о какой-то нестабильности. Очень может быть, что очень многие виды уже были подточены какими-то долговременными процессами и стояли на пути вымирания, а катастрофа просто ускорила процесс.
- С другой стороны, следует иметь в виду, что продолжительность периода вымирания не может быть точно оценена из-за эффекта Синьора — Липпса, связанного с неполнотой палеонтологических данных (время захоронения последнего найденного ископаемого может не соответствовать времени исчезновения таксона).
- Часть гипотез имеют недостаточно фактических подтверждений. Так, не найдено никаких следов того, что инверсии магнитного поля Земли влияют на биосферу; нет убедительных доказательств того, что маастрихтская регрессия уровня Мирового океана могла вызвать массовое вымирание таких масштабов; нет доказательств резких скачков температуры океана именно в этот период; также не доказано, что катастрофический вулканизм, в результате которого образовались деканские траппы, был повсеместным, или что его интенсивность была достаточной для глобальных изменений климата и биосферы.
Недостатки биосферной версии[править | править код]
В вышеприведённом виде версия использует гипотетические представления о физиологии и поведении динозавров, при этом не сопоставляя все изменения климата и течений, имевшие место в мезозое, в конце мелового периода, а потому и не объясняет одновременное вымирание динозавров на изолированных друг от друга материках[33].
- ↑ https://www.britannica.com/science/K-T-extinction
- ↑ 1 2 3 Алексей Симонович Кондрашов, к. б. н., Мичиганский университет, США. Продолжение дискуссии о «научной истине».
- ↑ Most comprehensive tree of life shows placental mammal diversity exploded after age of dinosaurs.
- ↑ Динозавры из палеогена // Палеонтологический портал «Аммонит.ру», 01.05.2009.
- ↑ Sheehan P. M. et al. Sudden extinction of the dinosaurs: latest Cretaceous, upper Great Plains (англ.) // Science. — 1991. — Vol. 254, no. 5033. — P. 835—839.
- ↑ Milner A. C. Timing and causes of vertebrate extinction across the Cretaceous-Tertiary boundary (англ.) // Geological Society, London, Special Publications. — 1998. — Vol. 140. — P. 247—257.
- ↑ Peter Schulte et al. The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary, Science, 05 Mar 2010: Vol. 327, Issue 5970, p. 1214—1218. doi:10.1126/science.1177265.
- ↑ Алексей Симонович Кондрашов, к. б. н., Мичиганский университет, США. Было бы здорово в Ханты-Мансийском Археопарке заменить бронзовых мамонтов на настоящих.
- ↑ Динозавры вымерли из-за падения астероида // Русская служба Би-би-си, 5 марта 2010 г.
- ↑ 1 2 David Tytell. Did a Comet Swarm Kill the Dinosaurs? // Sky & Telescope, 14 May 2004.
- ↑ См., напр., Keller, G. et al. Chicxulub impact predates the K-T boundary mass extinction // Proc. Nat. Academy of Sci. of the USA, vol. 101, no. 11, p. 3753—3758 (2004).
- ↑ Earth Impact Database.
- ↑ Любопытно отметить, что такая же гипотеза привлекается среди других для объяснения массового пермского вымирания.
- ↑ Dingus, Norell, 2011, p. 114.
- ↑ Своей особой популярностью среди широкой публики эта гипотеза не в последнюю очередь обязана тем, что её очень ярко и наглядно изобразили создатели известного телесериала «Прогулки с динозаврами» с помощью компьютерной графики.
- ↑ Kevin O. Pope, Kevin H. Baines, Adriana C. Ocampo, Boris A. Ivanov. Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact (англ.) // Journal of Geophysical Research[en]. — 1997. — Vol. 102, no. E9. — P. 21645—21664. — ISSN 0148-0227. — doi:10.1029/97JE01743. — PMID 11541145.
- ↑ 1 2 Charles G. Bardeen et al. On transient climate change at the Cretaceous−Paleogene boundary due to atmospheric soot injections, 17 July 2017 (received for review 30 May 2017).
- ↑ Julia Brugger et al. Baby, it’s cold outside: Climate model simulations of the effects of the asteroid impact at the end of the Cretaceous // Geophysical Research Letters. Volume 44, Issue 1, 16 January 2017, Pages 419—427, 13 January 2017.
- ↑ Жизнь вернулась в кратер Чиксулуб почти сразу после падения астероида. Элементы большой науки.
- ↑ Michael J. Henehan et al. Rapid ocean acidification and protracted Earth system recovery followed the end-Cretaceous Chicxulub impact, 2019.
- ↑ W. F. Bottke1, D. Vokrouhlický, D. Nesvorný. An asteroid breakup 160 Myr ago as the probable source of the K/T impactor // Nature, 2007, vol. 449, p. 48—53 doi:10.1038/nature06070.
- ↑ Chatterjee, Sankar. Multiple Impacts at the KT Boundary and the Death of the Dinosaurs (англ.) // 30th International Geological Congress : journal. — 1997. — August (vol. 26). — P. 31—54.
- ↑ Двойной удар // Космос-журнал.
- ↑ Рэндалл, 2016, с. 4.
- ↑ Keller G., Adatte T., Gardin S., Bartolini A., Bajpai S. Main Deccan volcanism phase ends near the K-T boundary: Evidence from the Krishna-Godavari Basin, SE India (англ.) // Earth and Planetary Science Letters (англ.)русск. : journal. — 2008. — Vol. 268. — P. 293—311. — doi:10.1016/j.epsl.2008.01.015.
- ↑ Dingus, Norell, 2011, pp. 115—116.
- ↑ 1 2 3 4 Кирилл Еськов, История Земли и жизни на ней: От хаоса до человека. Архивная копия от 14 июня 2008 на Wayback Machine — М.: НЦ ЭНАС, 2004. — 312 с. — 10 000 экз. ISBN 5-93196-477-0.
- ↑ А. Бурсм. Глава 9: Палеотемпературы и соотношения изотопов углерода по разрезу от кампана до палеоцена и граница мелового и третичного периодов в Атлантическом океане (неопр.) (недоступная ссылка). Дата обращения 14 июля 2009. Архивировано 2 сентября 2013 года.
- ↑ Matt Kaplan, Roland Sookias, Daryl Codron, Dinosaurs grew to outpace their young.
- ↑ State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact.
- ↑ Учёные узнали о прикончившем динозавров «двойном ударе».
- ↑ Динозавры вымерли из-за отсутствия самок. Архивная копия от 6 сентября 2010 на Wayback Machine. 21.04.2004, [1]. Архивная копия от 3 октября 2006 на Wayback Machine (англ.).
- ↑ Алексеев Александр Сергеевич. Массовые вымирания в фанерозое (рус.). — 1999.
- Brown, Barnum. A Triceratops Hunt in Pioneer Wyoming: The Journals of Barnum Brown & J.p. Sams : The University of Kansas Expedition of 1895. — High Plains Pr, 2004. — 188 p. — ISBN 0931271770.
- Лиза Рэндалл. Тёмная материя и динозавры: Удивительная взаимосвязь событий во Вселенной = Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the Universe. — М.: Альпина Нон-фикшн, 2016. — 506 p. — ISBN 978-5-91671-646-7.
- Dingus, Lowell; Norell, Mark. Barnum Brown: The Man Who Discovered Tyrannosaurus rex. — University of California Press, 2011. — 384 p. — ISBN 0520272617.
Почему вымерли динозавры | Наука
Земная кора хранит свидетельства множества катастроф. Мелово-палеогеновое вымирание, после которого 65 миллионов лет назад исчезли динозавры, плезиозавры и птерозавры, наиболее известно и изучено лучше всего. Тем не менее с этим событием по-прежнему связано немало загадок. Что послужило его основной причиной?
Старейшая и наиболее распространённая гипотеза связывает вымирание динозавров с падением астероида. Первоначально на данную мысль исследователей навело повышенное содержание элементов, не характерных для земной коры, в отложениях древностью 65 миллионов лет — именно тогда, как считается, вымерли динозавры. Позже катастрофу стали отождествлять с конкретным импактным событием — образованием кратера Чиксулуб на полуострове Юкатан (современная Мексика).

Обнаруженные в отложениях 65-миллионнолетней давности частицы сажи могут свидетельствовать о том, что падение астероида вызвало испарение и взрыв подземного нефтяного резервуара (худ. Donald E. Davis)
Способность десятикилометрового тела серьёзно напакостить в планетарных масштабах вызывала обоснованные сомнения. Но эти вопросы благополучно отпали после обнаружения на дне Индийского океана гигантского кратера, предположительно образованного астероидом 40 километров в поперечнике. Астероид, как и кратер, был назван Шивой. Затем были найдены ещё несколько воронок, оставленных меньшими, чем Чиксулуб, фрагментами Шивы.
Случившуюся тогда катастрофу проще описать, чем вообразить. Пронзив прикрытую плёнкой океана земную кору, Шива взорвался, выбив воронку глубиной 80 километров. Попробуйте представить трёхкилометровую толщу вод, водопадом летящую по склонам кратера, чтобы встретиться с кипящим камнем и обратиться в пар. Моря, выплёскивающиеся на берег трёхсотметровыми валами, чтобы опустошить миллионы квадратных километров суши. Небо — низкое, чёрное, непроницаемое, состоящее, кажется, только из пепла и пара. Главный же урон нанесли извержения, вызванные сотрясением земных недр, и отравляющие почву кислотные дожди. После падения Шивы Земля не могла успокоиться миллион лет!

После падения Шивы изливающаяся из трещин лава образовала в Индии деканские траппы — базальтовые поля толщиной два километра и площадью с Францию (Zina Deretsky)
Катаклизм, способный погубить всё живое, на первый взгляд исчерпывающе объясняет вымирание ящеров. Но гипотеза между тем имеет сразу две слабости. Во-первых, абсолютно непонятно, каким образом вышеописанные ужасы могут иметь отношение к делу. Динозавры начали вымирать задолго до падения Шивы, да и после него продолжали бороться за жизнь ещё несколько миллионов лет.
Во-вторых, даже если допустить, что падение астероида ускорило гибель гигантских ящеров, непонятно, почему в число пострадавших попали лишь динозавры, тогда как черепахам, крокодилам, змеям, птицам и млекопитающим Шива нисколько не навредил.
Ещё о динозаврах

18.10.2017
Могло ли так сложиться, что гигантские ящеры сохранились до наших дней? И каким бы стал тогда мир?
Альтернативной «космической» причиной вымирания мог послужить близкий взрыв сверхновой звезды, вследствие которого на поверхность планеты обрушились потоки смертоносного излучения. Однако эта гипотеза имеет те же изъяны, что и предыдущая. Кроме того, следы вспышки, способной уничтожить всё живое в радиусе 30 световых лет, современные телескопы со столь малого (по меркам астрономии) расстояния, скорее всего, обнаружили бы даже спустя 65 миллионов лет. Но в ближайших окрестностях Земли не найдено остатков сверхновых.
Впрочем, источником радиации не обязательно могла оказаться звезда, решившая завершить свой жизненный путь со спецэффектами и максимальным уроном для окружающих. Аналогичный эффект могло иметь, например, временное «отключение» магнитного поля планеты, защищающего биосферу от потоков космических частиц. По неизвестным причинам магнитное поле Земли время от времени действительно слабеет и меняет полярность, исчезая в момент «переключения» полюсов. Но лишь за последние 5 миллионов лет смена полярности происходила двадцать раз без каких-либо последствий для обитателей планеты.
Не раз звучала и чисто фантастическая гипотеза, что динозавров умышленно истребили пришельцы, дабы расчистить дорогу млекопитающим и приблизить появление человека. Если так, то представители сверхцивилизаций не разбираются в биологии. Ведь на эволюционном пути от примитивного насекомоядного к человеку разумному — то есть с дерева на землю, собирать камни и палки, — ни один динозавр не стоял.
Кого считают динозаврами?

Под названием «динозавры» объединяются два отряда теплокровных рептилий — птицетазовые и ящеротазовые. К птицетазовым относятся такие необычные ящеры, как утконосый игуанодон, рогатый трицератопс, вооружённый моргенштерном и оборудованный солнечной батареей стегозавр, а также броненосный анкилозавр. Все птицетазовые были крупными (от 1 до 10 тонн) травоядными. Характерным признаком отряда был роговой клюв.
Ящеротазовые динозавры делились на два подотряда: тероподы и зауроподы. К последним относились гигантские травоядные ящеры с длинной шеей — диплодоки, бронтозавры и прочие. Тераподы же («звероногие» ящеры) представляли собой двуногих хищников самого разного размера. Некоторые рептилии этого подотряда были не больше курицы, но в него же входили тираннозавр и спинозавр. Именно от этой, наиболее прогрессивной ветви динозавров, «изобретениями» которой стали перьевой покров и полые кости, произошли птицы.
Общий признак всех динозавров — ноги, «убранные» под корпус. У прочих рептилий конечности располагаются по бокам тела.
Ледниковый период?
Если искать причины вымирания динозавров на Земле, то наиболее очевидным вариантом кажется изменение климата. А климат на планете в ту пору как раз менялся. На протяжении почти всего мелового периода было удивительно тепло. Полярные шапки отсутствовали, и даже на севере современной Сибири условия напоминали средиземноморский курорт. Крокодилы в ту пору населяли реки до широты Архангельска. Динозавры же и млекопитающие встречались у самых полюсов.

Млекопитающие, жившие во времена динозавров, сами ещё не слишком отличались от рептилий. Температура тела ехидны колеблется от 28 до 30 градусов. Морозы зверёк переносить не способен
Холодать стало 70 миллионов лет назад. Но, во-первых, процесс шёл неспешно. В начале палеогена (66 млн лет назад) на севере Гренландии всё ещё росли лиственные леса. Во-вторых, появление ледниковых шапок лишь сместило пригодную для жизни зону к экватору. Теплолюбивые крокодилы просто переселились южнее, в территории, прежде необитаемые. Ведь в меловой период субтропическая, тропическая и экваториальная зоны представляли собой пустыню, раскалённую, как Долина Смерти, и сухую, как Атакама.
В любом случае, похолодание не давало преимуществ древним млекопитающим. Динозавров же не пугала даже полярная ночь. Небольшие хищные тероподы зимой скрывались в норах и впадали в спячку. Припорошенные снегом диплодоки просто цепенели, экономя тепло. Некоторые ящеры даже научились использовать жар горячих источников, чтобы согревать кладки яиц.

Мегазостродон — «саблезубая белка», жившая 200 миллионов лет назад
Полностью теплокровными динозавров, которые с грехом пополам поддерживали температуру тела на уровне 25 градусов, конечно, назвать было нельзя. Но то же самое касалось и примитивных млекопитающих.
Трудно возложить ответственность за вымирание и на изменение состава атмосферы, продолжавшееся на протяжении всего мелового периода. Концентрация кислорода в воздухе, изначально достигавшая 40–45%, постепенно снизилась до современного уровня. В конце периода (это и стало причиной похолодания) начала падать концентрация углекислоты, в эпоху ящеров вдесятеро более высокая, чем сейчас. Но изменения в атмосфере происходили крайне медленно. И непонятно, каким образом они могли бы затронуть интересы динозавров.

Молодые тираннозавры, которые, в отличие от взрослых «суперпадальщиков», передвигавшихся со скоростью 7 км/ч, были способны бегать и охотиться, долгое время считались отдельным видом теропод
Тем не менее жертвы были. В середине мелового периода вымерли ихтиозавры. При высокой концентрации кислорода лёгочное дыхание давало хладнокровным рептилиям бесспорное преимущество в сравнении с жабродышащими акулами. Но когда кислорода стало меньше, встал вопрос, нужны ли в природе рыбоящеры, если обыкновенные рыбы ничем им не уступают.
Кислород накапливался в течение юрского периода, ещё более пышного и обильного, чем меловой. Затем избыток этого газа был захоронен в форме грандиозных отложений углекислого кальция (давшего название геологическому периоду мела). Но откуда в атмосфере взялось столько лишнего углерода?

По одной из версий, причиной вымирания травоядных ящеров могли стать яды, которыми цветковые растения защищаются от врагов. Ведь в желудке крупного динозавра могло помещаться несколько центнеров пищи
Третья из «планетарных» гипотез объясняет гибель динозавров метановой катастрофой. Колоссальное количество углеводородов находится на Земле в форме гидратов — похожих на снег кристаллов, представляющих собой неустойчивые соединения природного газа и воды. Гидраты удерживаются в твёрдом состоянии благодаря давлению и низкой температуре — их залежи сосредоточены под вечной мерзлотой и донными отложениями океана. Согласно гипотезе «метангидратного ружья», повышение температуры моря может спровоцировать лавинообразный процесс выделения метана. Помимо усиления парникового эффекта, катастрофа чревата и серией взрывов, мощность которых придётся считать на гигатонны. Ведь молнии будут воспламенять воздушногазовую смесь.
Предполагается, что подобное событие вполне могло завершить эпоху динозавров. Однако у этой гипотезы есть крупный недостаток: гидратных залежей в меловой период существовать не могло. Ведь на протяжении мела Земля охлаждалась, а не нагревалась, парниковый эффект снижался, крошечные участки вечной мерзлоты были лишь в горах Антарктиды, а температура придонных вод на океанском дне достигала 20 градусов.
Впрочем, в определённом смысле метановая катастрофа тогда действительно произошла. «Ружьё» выстрелило. Древние запасы метана, а также новые порции газа, выделяющиеся при интенсивном формировании новых и «дозревании» старых угольных залежей, были выброшены в атмосферу. Но поступал и окислялся этот газ постепенно, на протяжении 80 миллионов лет.
Все «катастрофические» гипотезы обладают одним недостатком. Они не объясняют, почему вымерли строго определённые отряды рептилий. Разгадка исчезновения динозавров должна крыться в особенностях их биологии. И в гипотезах, объясняющих вымирание с этой точки зрения, недостатка нет.
Отмечено, например, что яйца крокодилов, отложенные в более суровых условиях, отличаются увеличенной толщиной скорлупы. К тому же температура песка, в который закапывается кладка, оказывает влияние на пол зародыша. Чем ниже температура, тем больше вылупится самцов. Так, может быть, похолодание привело к тому, что из яиц динозавров перестали вылупляться самки? Или все кладки разом погибли, потому что крошечные ящеры не могли взломать упрочнившуюся на холоде скорлупу?
Уязвимость таких гипотез заключается в том, что строятся они на основании наблюдений за крокодилами. Но крокодилы-то выжили, а значит, упомянутые свойства их яиц не могли сыграть роковую роль на рубеже мела и палеогена. И много ли общего между крокодилами и живородящими плезиозаврами или несущими яйца птеродактилями?

Лёгкий скелет понадобился динозаврам для использования своего наиболее ценного «изобретения» — бега. До динозавров, рискнувших оторвать передние конечности от земли, сухопутные животные передвигались только шагом
Рассматривается также версия губительной эпидемии. Данная гипотеза хорошо объясняет характер вымирания. Действительно, вирус будет выбирать жертвы со сходными ДНК. Тем не менее предположение выглядит фантастическим. До сих пор неизвестен микроорганизм, способный угрожать существованию хотя бы одного вида, не говоря уж о семействах и отрядах. Паразиту невыгодно быть слишком смертоносным. Ведь, уничтожая хозяина, он уничтожает и себя.
Несостоятельной представляется и гипотеза генетического вырождения. Конечно, 20-40-тонные диплодоки и бронтозавры не могли быть многочисленными и вели полустационарный образ жизни, делая буквально несколько шагов в день. Это могло бы приводить к систематическому близкородственному скрещиванию, если бы динозавры уже рождались огромными. Но вылупившийся из яйца диплодок был очень подвижным существом размером с небольшую собаку. Ничто не мешало ему пускаться в странствия, чтобы уже в зрелом возрасте «осесть» в сотнях километров от места появления на свет.

Расчёты показывают, что двигаться гигантские четвероногие ящеры могли со скоростью от 4 до 10 км/ч
Проще всего объяснить вымирание вида тем, что его вытеснил вид более приспособленный. Но динозавры, на первый взгляд, не могли потерпеть поражение в конкурентной борьбе, так как соперников в природе не имели. Млекопитающие тогда ещё не были готовы выступить в качестве хищников и крупных травоядных. Десять миллионов лет после вымирания динозавров наиболее привлекательные экологические ниши либо занимались уцелевшими рептилиями и нелетающими птицами, либо просто пустовали.
Конкуренцией можно объяснить лишь вымирание птеродактилей. Уже в середине мела их отовсюду изгнали птицы, и птеродактили всем отрядом сгрудились на прибрежных скалах. Но на этом, последнем рубеже летающие ящеры встали насмерть, продержавшись 40 миллионов лет.

Первыми по-настоящему теплокровными животными стали зубастые птицы (на рисунке — позднемеловой «пингвин» гесперорнис)
Час пробил, когда похолодание согнало «полутеплокровных» птерозавров с обледеневших побережий. Птиц же оно только стимулировало к поиску новых источников пищи. Быстро возникли виды, владеющие техникой посадки и взлёта с воды и даже, подобно современным пингвинам, променявшие способность к полёту на навыки подводного плавания. Птеродактили, которые умели целыми часами парить, почти не расходуя энергию, но, схватив добычу, вынуждены были добираться до берега вплавь, не имели шансов.
Чтобы динозавры вымерли, у них должно было быть некое общее слабое место. Им, видимо, оказались особенности размножения.

Динозавры, конечно, при случае поедали млекопитающих. Но не охотились на них систематически. Ведь звери, полагаясь на обоняние и слух, выходили на промысел ночью. А хищные рептилии, подобно птицам, не видели в темноте.
Поскольку скорлупа должна пропускать воздух, само яйцо не может быть слишком большим. Соответственно, детёныши динозавров вылуплялись совсем крошечными по сравнению со взрослыми. Кроме того, хоть наиболее сообразительные из ящеров и начали проявлять заботу о потомстве, охраняя кладки и молодь, выкармливать своих отпрысков им было нечем. Динозаврик, который не получал концентрированной пищи в виде молока и с первых дней существования добывал пропитание самостоятельно, рос медленно. Для достижения зрелости крупному ящеру требовалось несколько десятилетий.
Даже у самых продвинутых рептилий «детская смертность» оставалась колоссальной. И млекопитающие сумели воспользоваться этим обстоятельством. Ещё не бросая вызов взрослым ящерам, насекомоядные тем не менее конкурировали с молодью динозавров, вынужденной питаться жуками и ящерицами.

Плезиозавры, которые высматривали рыбу сверху, с высоты собственной шеи, и ловили добычу (в том числе плывущих домой птеродактилей) у самой поверхности, тоже не выдержали конкуренции с птицами (худ. Дмитрий Богданов)
Спусковым механизмом катастрофы стало, скорее всего, появление травы. Именно отсутствие травяного покрова отличало ландшафты мелового периода, украшенные, помимо деревьев, лишь кустами папоротников и пятнами мха, от современных. Зелёным ковром, создающим дёрн и удерживающим почву от выветривания и вымывания, Земля обзавелась 70 миллионов лет назад.
Под прикрытием зарослей травы, которые позволяли охотиться за личинками днём, да ещё и ограничивали обзор (что снизило роль зрения в охоте), первобытные ёжики перешли в решительное наступление. Чаша весов склонилась в пользу зверей.
Первыми — ещё за несколько миллионов лет до окончания мелового периода — пали небольшие хищные тероподы. В том числе самые прогрессивные из рептилий — теплокровные (судя по всему) велоцирапторы. И в образовавшуюся брешь ринулись полчища древних кроликов из отряда многобугорчатых.

Весивший всего 20 килограммов стремительный, хитрый и смертоносный велоцираптор охотился на мелких травоядных. Но эту нишу в меловой период занимала лишь молодь крупных ящеров
Тем же приёмом, сокращая доступные молодым динозаврам ресурсы, величественных диплодоков в конкурентной борьбе повергли мелкие зверьки, не отличающиеся ни умом, ни проворством. Но всю траву переесть было нелегко, и побоище на лугах, так и не завершившись в юре, продолжилось в палеогене.
Последними вымерли успевшие приспособиться к питанию травой трицератопсы и самые известные из ящеров — тираннозавры.
Когда вымерли динозавры? — Звезда Мидгарда — LiveJournal
Динозавры еще в 16 веке в войнах принимали участие. Очень многое не вписывается в историю, которую преподают нам в школах и не только. Мы свято верим, что динозавры вымерли миллионы лет назад, ведь это официальная версия, но так ли это на самом деле? Оказывается есть масса гипотез о том, что эти доисторические животные жили рядом с человеком еще долгие годы, уже после «Рождества Христова». Картина «Самоубийство Саула» художника Питера Брейгеля Старшего, 1562 года, тому прямое подтверждение. На ней, среди прочего войска, изображены всадники верхом на динозаврах! (Сергей Изофатов).
Давно появилась эта мысль (ниже постараюсь ее изложить). И, вот, попалась на глаза вполне научная информация об уцелевшей органике в костях динозавров. Согласитесь, за 65 млн. лет. любой органический материал разложится на минеральные вещества, либо окаменеет, приобретет так же неорганические признаки.Но, не смотря на такой возраст, существуют вот такие факты:
На протяжении двадцати лет исследователи недоумевают, обнаруживая в костях вымерших «миллионы лет» назад динозавров следы ДНК и радиоактивного углерода.
Многие окаменелости динозавров включают в себя фрагменты настоящих костей, которые не успели минерализоваться, иными словами окаменеть. Для многих исследователей содержимое этих костей оказалось полной неожиданностью. Начиная с 90-х годов прошлого века ученые сделали целый ряд открытий, обнаружив в костях динозавров клетки крови, гемоглобин, легко разрушаемые белки и фрагменты мягких тканей, в частности эластичных связок и кровеносных сосудов. И что заслуживает особого внимания – ДНК и радиоактивный углерод.
Эволюционистам теперь предстоит решить грандиозную проблему, чтобы объяснить предположительно 65-миллионный возраст этих костей. Как сказала доктор Мэри Швайцер, причастная к открытию кровяных клеток,
«Если проба крови изменяется до неузнаваемости уже через неделю, каким же образом эти клетки могли уцелеть?»
А действительно, каким? В организме, вымершем миллионы лет назад, они, конечно же, уцелеть бы не смогли. Они могли сохраниться только в останках, которые были быстро погребены при катастрофических условиях и находились под слоем осадочных пород. Что прекрасно объясняется глобальным потопом.
Но поскольку эволюционное мировоззрение занимает в научных кругах прочные позиции, опубликовать результаты такого исследования оказалось достаточно сложно. «Один рецензент сказал мне, что для него неважно, на что указывают данные, это просто невозможно» – рассказывает доктор Швайцер. «В ответном письме я спросила у него: «Тогда какие данные Вас убедят?» – «Никакие».
Швайцер вспоминает, как первоначально ее внимание привлек ярко выраженный трупный запах, исходивший от скелета тираннозавра, найденного в окрестностях Хелл-Крика, штат Монтана. Когда она упомянула это Джеку Хорнеру, палеонтологу со стажем, он ответил, что так пахнут все кости с Хелл-Крика. Убеждение в многомиллионном возрасте костей динозавров настолько глубоко сидит в сознании палеонтологов, что никто из них ни разу не обратил внимание на нетипичный «запах смерти» – прямо у них под носом. Даже сама Швайцер, несмотря на многие сделанные ею открытия, по всей видимости, не может или не хочет отойти от устоявшегося мировоззрения. Обратите внимание на хронологию открытий, сделанных за два десятилетия – на очевидные и последовательные указания на то, что что-то прогнило в палеонтологическом королевстве с его теориями о динозаврах, вымерших миллионы лет назад.
В 1993 г., неожиданно для себя Мэри Швайцер обнаруживает в костях динозавров клетки крови.
В 1997 г., обнаруживают гемоглобин, а также различимые кровяные клетки в костях тираннозавра.
В 2003 г., следы протеинового остеокальцина.В 2005 г., эластичные связки и кровеносные сосуды.
В 2007 г., коллаген (важный костный структурный белок) в кости тираннозавра.
В 2009 г., легко разрушаемые белки эластин и ламинин, и снова коллаген в утконосом динозавре. (Если бы останкам было действительно столько лет, сколько принято датировать, в них не было бы ни одного из этих белков).
В 2012 г., ученые сообщили об обнаружении клеток костной ткани (остеоцитов), белков актина и тубулина, а также ДНК(!). (Вычисленные по результатам исследований темпы распада этих белков и особенно ДНК указывают на то, что они не могли храниться в останках динозавра в течение, как предполагают, 65 миллионов лет после их вымирания)
В 2012 г., ученые сообщают об обнаружении радиоактивного углерода. (Учитывая насколько быстро распадается углерод-14, даже если бы останкам было сто тысяч лет, в них не должно было бы остаться и следа его присутствия!)
***
В Канаде на территории Парка динозавров ученым смогли обнаружить в костях динозавра мелового периода структуры, которые напоминают красные кровяные тельца и коллагеновые волокна. Находки позволяют по-новому взглянуть на устройство тела древних живых существ.
Для того, чтобы найти следы органики, клеток и прочих элементов динозавровой плоти исследователи придумали особый метод анализирования фотоснимков, которые получаются с помощью электронного и ионного микроскопов. Последний применяется в ИТ-индустрии при выискивании дефектов в чипах.
Таким образом, британцы сделали это удивительное открытие не вследствие обнаружения окаменелостей, а благодаря уникальному методу анализа останков динозавров, а также забытым вот уже как сто лет экспонатам из Музея естествознания в столице Великобритании.
Принято считать, что белковые молекулы распадаются быстро и сохраняются в окаменелостях не дольше четырех млн лет. После чего остаются фрагменты, которые не могут дать особых представлений о белковой структуре.
Ученый Сержио Бертацо вместе с коллегами, изучая плохо сохранившиеся кости древних рептилий, заметил довольно необычные яйцевидные образования с весьма плотным ядром. На ум сразу же пришли эритроциты.
Исследователи стали сравнивать их с капелькой крови ныне живущего страуса — в ионном масс-спектрометре они напоминали красные кровяные тельца страуса эму.
Ученые тут же схватились за аргумент, говорящий в пользу теплокровности вымерших динозавров.
В еще одном фрагменте кости обнаружились волокнистые структуры, похожие на спираль коллагеновых волокон. Так как устройство этого белка у различных групп животных различается, палеонтологи обзавелись возможностью сформулировать новый инструмент классификации рептилий.
Эксперты прибегали к нескольким аналитическим техникам. Местонахождение и состав мягких тканей в окаменелых остатках устанавливали при помощи электронного микроскопа. Далее ионным пучком лаборанты рассекали образцы и рассматривали их структуру.
«Теперь же нам нужны дальнейшие изыскания, так как мы хотим выяснить чем в действительности могут быть структуры, которые мы наблюдаем внутри костей динозавров. Однако мы считаем, что они сравнимы с красными кровяными клетками и волокнами коллагена. И если мы сможем подтвердить это, тогда в наших руках окажется новый способ окунуться в прошлое динозавров и осмыслить то, как они росли и развивались», подчеркнул Бертацо.
О своем открытии палеонтологи сообщили в журнале Nature Communications.
***
Ну, а сейчас предлагаю посмотреть, где и как находят кости динозавров.
Кладбища динозавров
Кладбища динозавров в Китае
Холм, который потревожили дорожные строители, нашли кости
В другом месте в Китае. Скелет покоится не на огромной глубине, как это должно быть. Ведь за 60 млн. лет уровень грунта над ним должен накопиться огромный (выпадение пыли и эрозия, которая приносит грунтовый материал)
Тоже небольшая глубина залегания
Вообще скелет на поверхности
В Китае находят яйца динозавров в окаменевшей глине
Мексика:
Археологи раскопали в Мексике самое большое на планете кладбище динозавров. На площади 200х50 всего было найдено 14 скелетов:
Судя по расположению этих костей – динозавр попал в «мясорубку». Источник
Кости в склоне холма
Парк динозавров в округе Альберта (Канада):
Такой возраст динозаврам дают еще по причено того, что их кости находят в склонах этих холмов:
У геологов есть данные по возрасту этих слоев. Ведь, они накапливались миллионы лет… А принять практически моментальный период образования слоев, как показано здесь http://sibved.livejournal.com/185060.html при катаклизме — это поему-то не принимают. Хотя некоторые научные круги принимают же гипотезу гибели динозавров при катаклизме – от падения астероида. Но развития и стройной модели она не получила.
Кладбища динозавтров находят на определенной широте. Скорее всего, им подходил только этот климат в этих широтах. Как и слонам в наше время нужна огромная кормовая база саванн – динозаврам с их размерами нужна была буйная растительность. Севернее из гигантов жили мамонты, шертистые носороги. И мое мнение – мамонты с динозаврами жили примерно в одно время. Погубил их один глобальный катаклизм с последствиями в виде гигантской волны и потопа. Возможно, это было не в поздние исторические времена, но человек в то время уже существовал.
Пустыня Гоби:
Кости практически на поверхности
Этот экземпляр как-будто ходил еще пару лет назад.
А этот плавал здесь недавно в геологическом времени.
Яйцо динозавра из Монголии
Разные виды динозавров погибали одновременно. Перед катастрофой все были одинаковы
Понятна ли моя мысль, что есть вероятность, что динозаврам, которых находят у поверхности – не 65 млн. лет?
И тогда становятся понятны мотивы камней Ики:
Коллекция Джульсруда:
Возможно, что это фэнтази из тех времен, а может быть и нет?
Динозавр на стене храма в Камбоджи:
И более современное:
Есть множество случаев, которые собирала в свое время криптозоология. Возможно, на западе кто-то до сих пор это делает. У нас в основном, этим занимаются такие энтузиасты, как группа Космопоиск.
Источники:
[Spoiler (click to open)]https://cont.ws/post/240462?_utl_t=vk
http://my.mail.ru/mail/zhans_1959/photo/19172
http://encyclopedia-of-dinosaursdinosaurs.blogspot.ru/2014/11/finds-in-north-america.html
Почему вымерли динозавры: обзор всех версий
Динозавры вымерли! Это, пожалуй, единственный факт о них, с которым согласны все учёные. А вот касательно причин исчезновения гигантских ящеров до сих пор ведутся споры. Популярным является мнение, что их массовая гибель была вызвана столкновением гигантского астероида с Землёй. Однако есть много других интересных предположений, которые могут дополнять общепринятую теорию или же рассматривают альтернативные взгляды. Сегодня мы поговорим о том, почему вымерли динозавры.
Когда произошло вымирание динозавров?
Следует отметить, что вымирание не было мгновенным, как это обычно нам преподносят некоторые фильмы и телепередачи. Даже если отталкиваться от теории столкновения Земли с астероидом, то после этого все динозавры не погибли сразу, но процесс уже был запущен…
Началось вымирание в конце так называемого «мелового периода» (около 250 млн. лет тому назад) и длилось около 5 миллионов лет (!). В этот период исчезло множество видов животных и растений.
Тем не менее, динозавры довольно долго были доминирующим видом на Земле – около 160 миллионов лет. В этот период исчезали и появлялись новые виды, динозавры эволюционировали, приспосабливались к изменениям климата и смогли пережить несколько массовых вымираний, пока не случилось нечто, что привело к постепенной и окончательной их гибели.
Для справки: «Человек разумный» живёт на Земле лишь 40 тыс. лет.
Кто пережил вымирание?
Изменения климата на Земле в меловый период поубавило разнообразие жизни, но потомки многих тогдашних видов сегодня радуют нас своим присутствием. К ним относятся крокодилы, черепахи, змеи и ящерицы.
Млекопитающие тоже сильно не пострадали и после полного исчезновения динозавров смогли занять доминирующее положение на планете.
Может сложиться впечатление, что гибель живых существ на Земле была избирательной, и что были сформированы именно те условия, в которых не смогли бы выжить динозавры. При этом остальные виды хоть и сильно пострадали, но существование могли продолжить. Эти мысли сильно будоражат умы почитателей различных теорий заговоров.
Кстати, слово «динозавр» с греческого языка дословно переводится, как «страшный ящер».
Версии вымирания динозавров
На сегодняшний день всё ещё доподлинно неизвестно что именно погубило динозавров. Гипотез много, а доказательств недостаточно. Начнём мы с версии с астероидом, которую очень популяризовали и во многом исказили СМИ и киномейкеры.
Астероид
В Мексике есть кратер Чиксулуб. Считается, что он образовался именно после падения того зловещего астероида, который спровоцировал массовое вымирание динозавров.
Как выглядело столкновение астероида с ЗемлёйНепосредственно сам астероид вызвал огромные разрушения в районе своего падения. Почти всё живое было в этой области уничтожено. А вот остальные жители Земли пострадали от последствий падения этого космического тела. По планете прошла мощная ударная волна, в атмосферу поднялись тучи пыли, проснулись спящие вулканы, планету окутали плотные облака, которые практически не пропускали солнечный свет. Соответственно в разы поубавилось количество растительности, которая была источником пищи для травоядных динозавров, а они в свою очередь позволяли выживать хищным ящерам.
Ещё много интересного в наших соцсетяхКстати, существует предположение, что в тот период на нашу планету упало два небесных тела. На дне Индийского океана найден кратер, появление которого датируется тем же временем.
Любители всё опровергать ставят под сомнение эту гипотезу. По их мнению, астероид не был столь крупным, чтобы запустить череду катаклизмов. К тому же и до этого события, и после – с землёй сталкивались другие похожие космические тела, но массовых вымираний они не провоцировали.
Версия, что этот астероид занёс на планету микроорганизмы, которые поразили динозавров, тоже имеет место, хоть и не столь вероятна.
Космическое излучение
Продолжая тему того, что именно космос убил всех динозавров, стоит рассмотреть и предположение, что к этому привел всплеск гамма-излучения недалеко от солнечной системы. Происходит это вследствие столкновения звёзд или взрыва сверхновой. Поток гамма-излучения повредил озоновый слой нашей планеты, что привело к изменению климата и мутациям.
Вулканическая активность
Мы уже упоминали, что астероид мог спровоцировать пробуждение спящих вулканов. Но это могло произойти и без его участия, а последствия всё равно были бы печальными.
Значительное усиление вулканической активности привело к тому, что пепел в атмосфере частично ограничил поступление солнечного света. А дальше – наступление вулканической зимы, уменьшение количества растений и изменение состава атмосферы.
Скептикам и в этом случае есть что сказать. Многие учённые считают, что изменения, вызванные аномальной вулканической активностью, были постепенными, а динозавры имели высокую способность к приспособлению, которая и помогала им переживать разливного рода капризы природы. Так почему они не могли адаптироваться в этот раз? Вопрос без ответа.
Резкое снижение уровня Мирового океана
Это понятие называют «маастрихтской регрессией». Единственная связь этого события с вымиранием динозавров прослеживается в том, что всё происходило примерно в один период. К тому же предыдущие большие вымирания иногда сопровождалось изменением уровня воды.
Возможно вас заинтересует: Что случится миром в следствии таяния ледяного покрова Антарктиды
Проблемы с пропитанием
Тут есть два варианта: либо вследствие изменения климата динозавры попросту не могли найти себе достаточно пропитания, либо появились растения, которые убили ящеров. Считается, что на Земле распространились цветковые растения, содержащие алкалоиды, которыми и отравились динозавры.
Смена магнитных полюсов
Такое явление периодически случается на нашей планете. Полюса меняются местами, а Земля на некоторое время остаётся без магнитного поля. Таким образом, вся биосфера становится беззащитной перед космическим излучением: организмы гибнут или мутируют. Причём всё может длиться на протяжении тысяч лет.
Дрейф материков и изменение климата
Эта гипотеза повествует о том, что динозавры по каким-то причинам не смогли пережить изменения климата, которые были вызваны дрейфом материков. Всё происходило довольно прозаично: температурные скачки, гибель растений, пересыхание рек и водоёмов. Очевидно, что движение тектонических плит сопровождалось и повышенной вулканической активностью. Бедные динозавры просто оказались неспособными к адаптации.
Расположение материков на конец мелового периодаИнтересно, что повышение температуры могло повлиять на формирование динозавров в яйце. В итоге могли вылупляться только детёныши одного пола. Подобное явление наблюдается и у современных крокодилов.
Эпидемия
Насекомые, сохранившиеся в янтаре, могут поведать учёным много интересного о древних временах. В частности удалось выяснить, что многие опасные инфекции начали появляться именно в период вымирания динозавров.
Мы уже знаем, что динозавры могли приспособиться к изменениям климата, но слаборазвитый иммунитет не мог защитить их от смертельной болезни.
Рекомендуем: С чем связано вымирание пчёл
Теория контролируемой эволюции
Сразу следует отметить, что эта теория пользуется популярностью в конспирологических кругах. Эти ребята считают, что некий иной разум использует нашу планету, как площадку для экспериментов. Вероятно этот «разум» на примере динозавров изучал особенности эволюции, но пришло время очищать экспериментальную площадку, дабы начать такие же исследования, но уже с млекопитающими в главной роли.
Таким образом, внеземной разум враз очищает Землю от динозавров и начинает новый этап эксперимента, основным объектом которого являемся мы – люди! Прям РЕН-ТВ какое-то. Но стоит признать, конспирологи умело всё преподносят и неплохо опровергают другие теории.
Динозавры против млекопитающих
Мелкие млекопитающие вполне могли уничтожить зубастых гигантов. Учёные не исключают жесткую конкуренцию между ними. Млекопитающие оказались более совершенными в плане выживания, им проще добывать пропитание и приспосабливаться к окружающей среде.
После динозавров наступила эра млекопитающихГлавным преимуществом млекопитающих стало отличие их способа размножения от способа размножения динозавров. Последние откладывали яйца, которые не всегда удавалось уберечь от тех же мелких зверьков. К тому же маленькому динозавру требовалось огромное количество пищи, чтобы вырасти до нужных размеров, а пропитание становилось добывать всё сложнее. Млекопитающие вынашивались в утробе, вскармливались материнским молоком, а дальнейшем не нуждались в слишком большом количестве пропитания. Тем более под носом были всегда яйца динозавров, которые можно было незаметно оприходовать.
Совпадение факторов
Многие учёные склонны полагать, что не следует зацикливаться на какой-то одной причине, ведь динозавры были очень живучими и за миллионы лет выдерживали немало сюрпризов со стороны природы. Скорее всему виной и изменение климата, и проблемы с пропитанием, и конкуренция с млекопитающими. Не исключено, что астероид стал своеобразным контрольным выстрелом. Всё это в совокупности сформировало именно те условия, в которых динозаврам не выжить.
Грозит ли вымирание людям?
Динозавры жили на Земле миллионы лет, люди – лишь несколько десятков тысяч. За этот сравнительно небольшой период мы смогли создать разумное общество. Но от вымирания это нас едва ли защит.
Версий исчезновения человечества довольно обширное количество, начиная от глобальных катастроф и эпидемий, и заканчивая всё той же космической угрозой в виде астероидов и взрывов звёзд. Впрочем, люди уже сегодня могут запросто прекратить своё существование – запасов ядерного оружия на Земле более чем достаточно для этих целей… Правда, часть людей всё же может спастись, если мы успеем колонизировать Марс или другую планету, подходящую для этих целей.
Заключение
Ответить на вопрос: «Почему вымерли динозавры?» сегодня с уверенностью нельзя. Все версии за неимением весомых доказательств существуют лишь на уровне предположений. Стоит обратить внимание, что динозавры вероятно впервые за миллионы лет оказались под воздействием нескольких перечисленных факторов, и в итоге уступили место млекопитающим.
как вымерли? Когда вымерли динозавры?
Динозавры – это древние существа, которые появились на планете примерно 225 миллионов лет назад. В течение 160 миллионов лет эти животные доминировали на планете. Период исчезновения занял около 5 миллионов лет, и вот уже примерно 65 миллионов лет в мире животных они отсутствуют. Существует много гипотез, почему исчезли динозавры. Как вымерли и прекратили своё существование эти животные, мы расскажем в нашей статье.

Появление динозавров
Планета Земля была заселена разными видами растений и животных 3 млрд лет назад. В процессе эволюции растения и животные появляются и исчезают, и каждый такой процесс имеет свой временной промежуток и период. Динозавры на планете жили в эру мезозоя – это триасовый, юрский и меловой периоды.
Первыми простейшими растениями были морские водоросли, а первыми животными – небольшие морские моллюски. Появление рыб произошло примерно 500 млн лет назад. Примерно 370 млн лет назад на сушу вышли первые животные – амфибии. Рептилии – это новая группа животных, которая появилась примерно 300 млн лет назад. У животных была чешуйчатая кожа, они могли откладывать яйца и постоянно находиться на суше. Следующими в цепочке эволюции стали динозавры. Вымерший вид животных дал толчок в развитии такой науки, как палеонтология.

Описание динозавров
Одними из удивительных животных, которые жили на планете, являются динозавры. Как вымерли и как жили эти большие животные, можно судить только по окаменевшим останкам. По ископаемым останкам можно сделать вывод, что они были рептилиями, как крокодилы, ящерицы, черепахи и змеи. Размер динозавров изменяется в большом диапазоне – от крошечных до гигантов. Обладали они четырьмя конечностями и хвостом. Стояли и передвигались динозавры на прямых конечностях, одни — на задних лапах, другие — на всех четырёх, третьи могли перемещаться как на двух, так и на четырёх конечностях. Многие динозавры обладали длинными шеями и зубами. Ареал обитания их был значительный, но 65 тысяч лет назад они внезапно вымерли.
Динозавры подразделяются на две группы: ящеротазовые и птицетазовые. Отличие групп состоит в строении костей таза. У ящеротазовых динозавров строение таза четырёхлучевое, а у птицетазовых – трёхлучевое. У некоторых видов птицетазовых были рога, шипы, панцири.

Появление интереса к динозаврам
В 30-х годах XIX века впервые были обнаружены окаменевшие останки динозавров. Тогда археологи не придали им особого значения и лишь спустя какое-то время стало понятно, что эти окаменелости принадлежат древним животным. Само понятие «динозавр» было введено английским зоологом Ричардом Оуэном в середине XIX века. С латинского языка «динозавр» переводится как «страшный», «опасный», «ужасный», а с древнегреческого языка – «ящер», «ящерица». С тех пор интерес к этим животным постоянно растёт. Сколько лет назад вымерли динозавры? Ответ на этот вопрос даёт наука палеонтология. Древних животных изучают учёные, снимают в фильмах, они становятся героями книг. И несмотря на такой интерес, на вопрос, отчего вымерли динозавры, точного ответа нет.
Эпоха динозавров
В конце пермского периода произошло образование единого материка – Пангеи. Характерной особенностью этого времени была глобальная вулканическая активность и исчезновение около 90 % животных. Лучше всех к новым условиям адаптировались рептилии. В начале триаса появилась группа рептилий под названием «пеликозавры». К середине триасового периода их сменила группа рептилий с названием «терапсиды». Параллельно с терапсидами развивалась новая группа рептилий – архозавры. Эта группа рептилий является предком всех динозавров, плиозавров, крокодиломорфов, ихтиозавров, плакодонтов и птерозавров. Следующий вид рептилий именовался текодонтами и был адаптирован к жизни на суше. И уже от них получили своё развитие динозавры. Вымершие животные хорошо приспособились и заняли доминантные позиции на суше, в воде и в воздухе.
В период триаса существовали следующие виды динозавров: целофизис, мусзавр и прокомпсогнат. Развивались и эволюционировали растительные динозавры.
Самые крупные животные жили в юрском периоде. В позднеюрском периоде стали появляться сухопутные животные – брахиозавр, диплодок и др.
В меловом периоде в морях и океанах стали преобладать хищные пресмыкающиеся. Появляются новые виды динозавров.

Окончание эпохи
Меловой период – это период расцвета гигантских ящеров, воздушных птеродактелей и морских рептилий. В конце мелового периода происходит раскол континента Пангеи на Гондвану и Лавразию. Климат на Земле становится значительно холоднее, на полюсах образуются ледяные шапки. Появляются цветковые растения, и увеличивается количество насекомых.
Всё это привело к вымиранию многих видов растений и животных, в том числе и динозавров. Вымерли они не в одночасье, но если учесть, что их господство длилось 160 млн лет, то их исчезновение произошло довольно быстро. Причины катастрофы, произошедшей в меловой период, до сих пор не ясны.
Но все ли динозавры вымерли? Потомками древних рептилий являются существующие сегодня крокодилы, ящерицы и птицы. Первые птицы появились в эпоху мела, и к концу эпохи у них уже было развитое оперение. Когда вымерли динозавры, птицы переняли эстафетную палочку эволюции.
Астрофизические гипотезы вымирания
Падение астероида является одной из распространённых версий. Время его падения совпадает с образованием кратера Чиксулуб (Мексика, полуостров Юкатан). Эти события произошли примерно 65 млн лет назад, в период, когда вымерли динозавры. Возможно, падение астероида повлекло за собой разрушительные действия, вследствие чего произошло массовое вымирание всего живого.
Гипотеза многократного падения гласит, что падение астероида происходило несколько раз. Кроме кратера Чиксулуб существует кратер Шива в Индийском океане, который образовался примерно в то же время. Эта гипотеза объясняет, почему вымирание происходило постепенно.
Ещё существует версия взрыва сверхновой звезды и столкновения кометы с Землей.

Геологические и климатические гипотезы вымирания
На планете происходили значительные изменения в период, когда начали исчезать динозавры. Как вымерли животные, предполагает теория изменения среднегодовых и сезонных температур. Крупным особям нужен тёплый и ровный климат. Вулканическая активность могла повлечь за собой изменение состава атмосферы и вызвать парниковый эффект. Большой выброс вулканического пепла мог спровоцировать вулканическую зиму, тем самым изменить освещённость Земли. Значительное понижение уровня моря, охлаждение океана, изменение состава морской воды и резкий скачок магнитного поля Земли так же могли способствовать вымиранию динозавров.
Эволюционно-биологические гипотезы вымирания
Одна из гипотез этой группы придерживается ситуации возникновения массовой эпидемии. Не исключено, что динозавры не смогли приспособиться к изменившейся растительности, что повлекло отравления. Велика вероятность уничтожения яиц и детёнышей первыми хищными млекопитающими. Ещё существует версия, что во время ледникового периода исчезли женские особи. Учёные предложили ещё одну версию гибели динозавров – удушье: в атмосфере произошло резкое уменьшение количества кислорода.
Почему исчезли динозавры?
Почему исчезли динозавры? Как вымерли эти древние животные? Ответы на эти вопросы дают разнообразные теории и гипотезы, но ни одна из них не отвечает на все вопросы в полной мере. Известно, что вымирание видов началось задолго до момента катастрофы, и астрономическая гипотеза в этом случае сомнительна. У многих теорий отсутствуют фактические данные, как, например, у гипотезы регрессии Мирового океана или изменения магнитного поля. Также отсутствие полноты палеонтологических данных может давать искажённую картину.
Комбинирование гипотез формирует более наглядную картину. Гипотезы, дополняя друг друга, дают ответы на большее количество вопросов, и картина того времени выглядит более прорисованной и детальной.
Процесс эволюции – вымирание старого и образование нового – последовательный. И процесс эволюции динозавров до конца мелового периода происходил естественно. Но по какой-то причине в конце мелового периода старые виды вымирали, а новые не появлялись, и, как следствие, произошло полное вымирание этого вида.

С точки зрения палеонтологии
Версия великого вымирания основана на следующих фактах:
- Появление цветковых растений.
- Постепенное изменение климата, вызванное дрейфом материков.
По мнению научного мира, наблюдалась следующая картина. Развитая корневая система цветковых растений, их лучшая приспособляемость к почвам быстро вытеснили другие виды растительности. Стали появляться насекомые, которые питались цветковыми растениями, а ранее появившиеся насекомые начали исчезать.
Корневая система цветковых стала разрастаться и препятствовать процессу эрозии почв. Поверхность суши перестала размываться, и в океаны перестал поступать питательный материал. Это привело к обеднению океана и гибели водорослей, которые, в свою очередь, являются производителями биомассы в океане. В воде произошло нарушение экосистемы, что стало причиной массового вымирания. Считается, что летающие ящеры тесно связаны с морем, поэтому цепочка вымирания перекинулась и на них. На суше они пытались приспособиться к зелёной массе. Стали появляться мелкие млекопитающие и хищники малого размера. Это являлось угрозой для потомства динозавров, так как яйца и детёныши динозавров стали питанием для появившихся хищников. В результате создались условия, отрицательные для возникновения новых видов.
Когда вымерли динозавры, эра мезозоя закончилась, с ней же закончилась и активная тектоническая, климатическая и эволюционная деятельность.

Дети и динозавры
Интерес к древним животным есть не только у взрослых, но и у детей. Сегодня проект «Почему вымерли динозавры?» включен в программу детских садов и начальных классов. Уникальность подобной деятельности заключается в том, что ребёнок самостоятельно развивает познавательные способности, ищет ответы на вопросы и получает новые знания. Вопрос о том, почему вымерли динозавры, для детей так же любопытен, как и для учёных. Интерес в первую очередь вызван тем, что этих животных сегодня на земле нет и точный ответ на вопрос о причинах их исчезновения до сих пор не получен.
Когда вымерли динозавры, или обман на миллионы (лет) ~ Проза (Очерк)

В пустыне Такла-Макан (Синьцзянь-Уйгурский автономный район Китая) издавна находят яйца гигантских доисторических ящеров. И не только находят. Местные жители, не избалованные щедротами своего засушливого края, научились использовать их в качестве дополнения к своему рациону.
Перед употреблением в пищу затвердевшее динозавровое яйцо проходит длительный процесс обработки: его вымачивают в специальном растворе, отчего сходит скорлупа и само яйцо размягчается, затем долго варят, засаливают и лишь потом с разными приправами подают к столу. Называется это самое, наверное, экзотическое на Земле блюдо «сымажо».
Однако не в его экзотичности дело. А в том, что яйца динозавров действительно вполне съедобны. И этот факт опровергает все здание – такое стройное и законченное… казалось бы — не только современной палеонтологии, но и прочих сопутствующих ей наук: геологии, биологии, истории, наконец.
Почему? Да потому что динозавры вымерли, как нам всем прекрасно известно – по уверениям этой самой палеонтологии – 65 миллионов лет назад, и все, что мы о них знаем, проистекает из их окаменелых останков, извлеченных из недр земли. Но что такое эти окаменелые останки? Это не просто кости или яйца, из-за своей древности ставшие твердыми, как камень; нет, это всего лишь копия этих самых костей и яиц, возникшая в результате процесса, известного как фессализация. Фессализация, или окаменение – это замещение кальция, содержащегося в живом организме, на кремний. В фессализированных ископаемых не остается ничего органического, что и позволяет им сохраняться на протяжении столь длительного периода времени до наших дней. Но… камень, пусть даже и имеющий форму яйца, остается камнем, и как бы вы его не вымачивали, размягчить его у вас не получится. Значит, «сымажо» изготовлено не из окаменевших, а всего лишь высохших яиц динозавров! А это настоящая катастрофа для палеонтологии, подрывающая все ее основы, и прежде всего – палеонтологическую летопись Земли!
Проблема с «неокаменевшими» яйцами динозавров состоит в том, что за 65 миллионов – да и просто за миллион – лет сохраниться они не могли физически. Потому что, потеряв свою влагу, яйца за этот срок сначала бы мумифицировались – совсем как египетские фараоны – а затем рассыпались бы в пыль. Но… они всего лишь высохли! И выходит, что цератозавры, диплодоки и прочие обитатели Такла-Макан отложили их не 65 миллионов лет, а буквально «вчера», каких-нибудь 10-12 тысяч лет назад! Кстати, такой же возраст имеют и останки других вымерших существ – мамонтов, чье замороженное в вечной мерзлоте мясо население севера еще совсем недавно также употребляло в пищу.
Но… динозавры – современники мамонтов?! Ведь 65 миллионов лет взяты не с потолка, все палеонтологи мира в один голос уверяют нас, что именно такая прорва времени прошла с тех пор, как гиганты кайнозойской эры сотрясали своими шагами землю. И не верить им нет никаких оснований!
На первый взгляд – да. А если разобраться и взглянуть на эти уверения непредвзято?
Уже на второй взгляд с удивлением понимаешь, что именно с потолка, и ниоткуда еще, и взяты эти самые пресловутые миллионолетия. А точнее – из безапелляционного заявления Чарльза Лайеля (1797-1875), основоположника современной геологии, современника и соратника другого широко известного Чарльза – Дарвина. Захваченный идеями последнего, Лайель сказал: «силы, ныне действующие как на земной поверхности, так и под нею, могут быть тождественны по роду и степени с теми, которые в отдаленные эпохи производили геологические изменения», а «геологи так неправильно истолковали признаки последовательности событий, что считали столетиями там, где цифры означали тысячелетия, и тысячелетиями там, где язык природы означал миллионы лет…» Сказал он это в ответ на нападки оппонентов теории эволюции, заявлявших, что на изменения, отмеченные в этой теории, времени существования Земли просто недостаточно. И… ему поверили! Без доказательств и фактов, просто – поверили! С тех пор палеонтологи и следуют указанным им курсом, не в силах сойти с проложенной колеи.
А как же все эти суперсовременные методы определения возраста ископаемых предметов, которым те же палеонтологи молятся, как иконам: радиоуглеродный метод, калий-аргонный метод, урановый метод? К сожалению, все они не только не отличаются хоть сколько-нибудь приемлемой точностью – например, ошибки, выдаваемые радиоуглеродным методом датировки, сопоставимы с возрастом исследуемого предмета – но и… протяженностью во времени, на которую их можно применять. Ведь все эти методы основаны на периодах полураспада тех или иных радиоактивных элементов, и, следовательно, не могут «увидеть» предмет, чей возраст превышает этот период более чем вдвое. Для урана, например – самого «долгоиграющего» из элементов – этот период составляет 24 миллиона лет; следовательно, окаменелости и предметы, которые старше 48 миллионов лет, ему уже неподотчетны. И снова мы возвращаемся к тому вопросу, с которого начали: откуда взялись 65 миллионов лет с момента гибели динозавров, не говоря уже о 5,5 миллиардах лет с момента возникновения Земли? И ответ, к сожалению, тоже остается неизменным: с потолка! Потому что палеонтологам это ВЫГОДНО!
Правда, справедливости ради, стоит заметить, что и сами палеонтологи, похоже, не слишком удовлетворены сложившимся в их науке положением и потому ищут другие, более подходящие, способы определения возраста. И даже нашли один такой – по скорости осадконакопления. Давно известно, что наша Земля «толстеет» — прибавляет в среднем по одному сантиметру за сто лет к своему диаметру. Происходит это за счет разложения органических останков – животных и растений – а также неорганических наносов: песка и глины, которые «поставляют» разрушающиеся из-за выветривания горы. Скорость этого накопления осадков, конечно же, неравномерна и зависит от местности – быстрее всего «толстеют» тропические джунгли, а медленнее всего – пустыни. И 1 сантиметр – это именно среднее значение этой скорости, в реальности же она составляет от 1 миллиметра до 3 сантиметров за век.
Прекрасно! Хоть какая-то определенность! Но… Ах, это пресловутое «но»! Как же оно все портит! Ведь так просто поверить, что ученые, занимающиеся всю свою жизнь своей наукой, знают, что делают и о чем говорят! Поверить – и не проверять их утверждения! А стоило бы! Да и к чему ждать? Давайте сделаем это прямо сейчас, тем более что больших усилий это не потребует. Ибо проверка эта доступна любому первокласснику. Итак, 1мм × 650 000 веков = 650 000 мм = 650м! Шестьсот пятьдесят метров! Именно столько осадков должно было накопиться за 65 миллионов лет, прошедших – якобы – с момента гибели динозавров!
Конечно, под давлением верхних слоев нижние прессуются, становясь чем глубже, тем тоньше, однако этой деформацией можно пренебречь, особенно если взять для расчета нижнюю планку – что мы и сделали. Если же произвести расчет по среднему значению осадконакопления, то мы получим фантастическую цифру в 6,5 километров! А осадочные породы, возникшие — якобы — во время возникновения Земли, должны залегать вообще на глубине… от 55 до 550 километров! Для сравнения – сверхглубокая скважина на Кольском полуострове, и по сей день остающаяся рекордной, едва дотянула до 13 километров.
Никто и никогда не производил раскопок на такой глубине! Да этого и не требовалось – кости динозавров находятся куда ближе к поверхности – два-три метра; в некоторых местах, как в том же самом Такла-Макане, они вообще едва прикрыты тонким слоем песка. Три метра… 3000 миллиметров… Рассчитаем возраст. 3000мм × 100 лет = 300 000 лет. Триста тысяч! Но не 65 миллионов! Впрочем, на такой глубине сжатие и спрессовывание словев почвы минимально, и вести расчет их возраста в данном случае можно именно что по среднему значению. А такой расчет дает и вовсе «детскую» цифру — 30 000 лет! Это — РЕАЛЬНЫЙ возраст пластов, залегающих на глубине 3 метров и содержащих кости «вымерших миллионы лет назад» динозавров. И, глядя на эти цифры, так и хочется закричать: люди! Проснитесь! Вас беззастенчиво, беспардонно, бессовестно обманывают! И динозавры – вовсе не дела давно минувших лет, а практически ваши современники! О чем прямо заявляют следы, обнаруживаемые теми самыми палеонтологами по всему миру. Окаменевшие следы динозавров – и людей, прошедших рядом с этими «ужасными» ящерами!
Впрочем, если принять время вымирания динозавров именно за тысячи, а не миллионы, лет, то такое соседство становится вполне объяснимым и даже закономерным. Ведь именно 30 тысяч лет назад человек начал обживать не только Европу, но и Америку, где в своем большинстве и обнаруживаются его «аномально древние» окаменевшие следы, мозоля глаза современным ортодоксам от палеонтологии.
Похоже, и сами палеонтологи уже не слишком удовлетворены классической хронологией. Во всяком случае, обнаружив в «окаменелой» ноге тираннозавра, откопанной в 1990 году в фомации Хелл-Крик в штате Монтана (США) мягкие ткани, гибкие (!), разветвлённые кровеносные сосуды с высохшим, но не разрушенным (!) гемоглобином и жилистую, но эластичную костную матрицу, автор находки, Мэри Хигби Швейцер, палеонтолог из Университета штата Северная Каролина, в статье в журнале «Earth» в 1997 г. «проговорилась»:
«Возможно, таинственные структуры были, в лучшем случае, производными крови, модифицированной тысячелетиями геологических процессов». («Perhaps the mysterious structures were, at best, derived from blood, modified over the millennia by geological processes»).
«Millennia» написала (тысячелетия), а не «millions», тем более не «tens millions» (десятки миллионов)!
Однако, как я уже отмечал ранее, 2-3 метра вовсе не универсальная глубина залегания окаменевших останков динозавров. В большинстве мест эти кости встречаются практически на поверхности, и возраст их в этом случае ограничивается «всего» несколькими тысячами, а порой и несколькими сотнями лет! А это уже исторический период, период, в который человечество, овладевшее письменностью, во всю использовало ее, занося все мало-мальски значимые события в свои летописи. Почитаем их?
По свидетельству древнекельтских хроник, король Моридд был убит и проглочен в 336 году до Рождества Христова гигантским монстром Белуа. Чудовище «проглотило тело Морвидуса (имя Моридда в латинской транскрипции. Авт.), как крупная рыба глотает мелкую».
Раннебритонскому королю Передару повезло больше — он победил в схватке с подобным чудовищем в местности Ллайн Ллайон (Уэльс). Британские хроники также рассказывают о множестве мест на территории нынешнего Уэльса, которые населяли чудовища Афанк и Каррог. Один из последних афанков был убит в 1693 году (!) Эдвардом Ллойдом в Ллайнар-Афанке на реке Конвей. А в хрониках Кентерберийского храма (Великобритания) отмечено, что в пятницу 16 сентября 1449 года неподалеку от деревни Литтл Конрад на границе графств Саффолк и Эссекс многие жители наблюдали схватку двух гигантских рептилий.
Однако почему же все так изменилось? Почему динозавры вымерли? Ну, учитывая все вышеприведенные летописи, а также неприведенные, но хорошо всем известные сказки, былины, легенды и мифы, можно смело утверждать, что последних представителей этого вида уничтожил, как это ни прискорбно, сам человек. Но динозавры все-таки были — и в этом палеонтология не врет — едва ли не самым многочисленным видом живых существ на нашей планете, и человек просто физически не мог послужить основной причиной их исчезновения. А значит, была другая, куда более весомая причина, и направившая «ужасных ящеров» на стезю вымирания. Человек же только завершил начатое ею. «Причина – юкатанский метеорит!» — скажет сейчас любой мало-мальски образованный человек. И… опять ошибется!
До 1991 года теорий, призванных объяснить необъяснимое исчезновение древних ящеров, существовало несколько десятков – от климатических изменений до взрыва сверхновой в окрестностях Солнца. Но потом был открыт Юкатанский метеоритный кратер, и об этих теориях забыли, решив, что разгадка наконец-то найдена.
Как ни парадоксально, но главный удар по этой «разгадке» наносят сами палеонтологи. Точнее, их находки – те самые окаменевшие следы, о которых говорилось чуть раньше. Причем человеческие следы здесь можно со спокойной совестью оставить «за кадром» и сосредоточиться именно на следах динозавров. Потому что если фессализация костных останков еще понятна, то вот фессализация следов никакого разумного объяснения не имеет – ни в свете метеоритной, ни какой другой теории. И палеонтологи, прекрасно отдающие себе в этом отчет, стараются лишний раз не привлекать к этим отпечаткам внимания: «Вот следы динозавра, прошедшего здесь 65 миллионов лет назад… А вот его скелет! Вы только посмотрите, какой он большой и страшный, как хорошо сохранились все его кости! А какие у него зубы! Такими зубами можно с легкостью перекусить легковую машину!.. Что, следы? А что следы? Следы как следы, что в них особенного? Не отвлекайтесь и смотрите лучше на кости!»
А следы-то как раз и являются особенными, потому что просто НЕ ДОЛЖНЫ существовать! Задумайтесь – для их консервации на МИЛЛИОНЫ (!) лет должны были (на те же миллионы лет): а) перестать идти дожди; б) дуть ветры; в) исчезнуть животные, способные их затоптать. На Земле существуют миллиарды живых существ, но нигде и никогда, за исключением «динозаврового» периода, их следы не сохранялись дольше недели; обычно этот срок равняется одному, максимум двум дням. И всё! Их стирают стихии и другие животные, прошедшие позже. А следы динозавров есть! Так что же могло послужить причиной их сохранения?
Несомненно, это была катастрофа глобального масштаба, и юкатанский метеорит не имеет к ней никакого отношения – именно из-за слабости своего воздействия на земную био- и геосферу. Его влияние даже по самым смелым предположениям астрономов, геологов и самих палеонтологов сходило на нет уже в радиусе 2000 километров от места его падения; где уж ему было повлиять на ВСЮ Землю? Причем – одновременно?
Так что же погубило древних ящеров?
Взрыв сверхновой можно отвергнуть сразу: рентгеновское излучение – единственное последствие такого взрыва, которое могло бы ощущаться на Земле – попросту стерилизовало бы всю нашу планету, уничтожив все живое вплоть до бактерий, так что сейчас мы с вами этот вопрос не обсуждали бы. Изменение климата? Сейчас доказано, что динозавры, по крайней мере, некоторые из них, были теплокровными, и даже резкое похолодание не смогло бы привести к их повсеместной гибели. Куда вернее оно уничтожило бы черепах и крокодилов, которые были современниками динозавров – и спокойно пережили их гибель. Тогда что?
В 1971 году в пустыне Гоби (Монголия) палеонтологи обнаружили окаменевшие останки протоцератопса и велоцераптора, сплетенные в схватке. Оба – и хищник, и его жертва – погибли СТОЯ, даже не разжав челюстей, стиснутых друг на друге. По всему выходило, что их смерть была неожиданной для них и одновременной, к тому же молниеносной. Что-то убило их на месте, превратив в памятники самим себе; и после этого они не только не рухнули бездыханными на землю, но к ним также не прикоснулся почему-то не один падальщик, оставив трупы нетронутыми.
Единственной причиной, по которой тело как бы застывает в той позе, что предшествовала его смерти, является… нет, не холод – даже жидкий азот не способен моментально заморозить живой организм, тем более столь упитанный, как протоцератопс, который по размерам и строению был очень похож на современную свинью – а… жар! Обжигающий жар, не менее 5000 градусов, приводящий к судорожному сокращению связок и сухожилий и мгновенно высушивающий их, в результате чего тело теряет всякую подвижность на очень долгое время. Однако этот жар должен действовать на тело не более нескольких секунд, иначе оно попросту сгорит, оставив только кучку обугленных костей. Одновременно такой жар и такой же продолжительности приводит и к спеканию почвы в некое подобие бетона, на котором прекрасно сохраняются следы тех существ, которые по этой почве прошли незадолго до воздействия на нее высокой температуры.
Ядерный взрыв? Или, учитывая масштабы произошедшего – глобальная ядерная война? Нет, хоть это и разочарует приверженцев погибших древних цивилизаций. Дело в том, что ядерный конфликт, как и взрыв сверхновой, уничтожил бы не только динозавров, но вообще все живое, и наша Земля представляла бы собой сейчас покрытый пеплом радиоактивный шар, пустой и совершенно безжизненный. Тогда что же?
Всё тепло и вся энергия, которую мы имеем на Земле, имеет один-единственный источник – Солнце. И именно на него мы должны обратить свое внимание в поисках ответа.
Давно известны 11-летние периоды активности нашего центрального светила. Однако были выявлены и другие циклы – вековые (продолжительностью 80-90 лет) и тысячелетние, длящиеся 1800-2000 лет. С последними, кстати, связывают возникновение пустыни Сахары, каких-то 4 000 лет назад являвшейся краем, полным жизни, воды и растительности. Удивительно, но все «солнца» древних майя, на которые они делили свой календарь, равнялись приблизительно 4 000 годам, за исключением последнего, пятого «солнца», длящегося 5126 лет. Впрочем, о майя разговор особый, а пока вернемся все-таки к нашему, не календарному, а самому что ни на есть настоящему Солнцу.
Итак, приблизительно каждые одиннадцать, 85 и 1900 лет с нашим светилом что-то происходит, и оно начинает вести себя «неадекватно»: вспыхивает, обрушивая на Землю настоящий шквал всевозможных излучений — рентгеновского, ультрафиолетового, инфракрасного, последствия чего ощущают на себе все метеозависимые – и не очень – люди. Даже самоубийство китов и дельфинов, выбрасывающихся вдруг на берег, связывают сейчас именно с этими вспышками, нарушающими навигацию морских млекопитающих в их родной стихии.
Мы уже настолько привыкли слышать об этих солнечных бурях, минимумах и максимумах солнечной активности, что не задумываемся над тем, что скрывается за этими, такими привычными, словами: что Солнце пульсирует и периодически взрывается, как сверхновая в миниатюре, сбрасывая в окружающее пространство излишки своей энергии. А значит, на нем могут происходить и куда более значительные флуктуации: взрывы, чья энергия в миллионы раз превышает энергию известных нам вспышек. Они МОГУТ происходить – и происходили! В результате одного из таких взрывов, произошедшего около миллиона лет назад (ох, уж эти миллионы!), у животных и птиц – и у людей – образовался пигмент меланин, подаривший нам черных воронов, черных кошек – и негроидную расу. Это – неоспоримый факт, признаваемый и астрономами, и биологами, и палеонтологами. Однако… давность произошедшего делает почему-то его в их глазах чем-то несущественным, и они отмахиваются от мысли, что подобное может – и должно! – повториться! И что одна из таких вспышек-взрывов стала причиной, по которой Земля на несколько секунд превратилась вдруг в раскаленную сталеплавильную печь, погубив динозавров и законсервировав их следы!
Так что не астероиды или пришельцы являются для нас главной угрозой, а наше солнышко, подарившее нам жизнь, но способное ее с легкостью отнять. Недаром, наверное, майя все-таки делили свою историю именно на «солнца»; похоже, они, в отличие от нас, прекрасно знали, что именно Солнце является началом всего сущего. И – его концом.